| Issue |
Aquat. Living Resour.
Volume 37, 2024
|
|
|---|---|---|
| Article Number | 7 | |
| Number of page(s) | 15 | |
| DOI | https://doi.org/10.1051/alr/2024005 | |
| Published online | 24 May 2024 | |
Research Article
Identification of critical essential habitat for demersal fish in the Gulf of Guinea
1
Unité Mixte de Recherche Science Agronomique et Génie Rural, Institut National Polytechnique Felix Houphouët Boigny, Yamoussoukro, Côte d’Ivoire
2
Laboratoire d’Evaluation des Ressources Vivantes et Aquatiques, Institut Mauritanien de Recherches Océanographiques et des Pêches, Nouadhibou, Mauritanie
3
Département Ressources Aquatiques Vivantes, Centre de Recherche Océanologiques, Treichville, Côte d’Ivoire
4
UMR DECOD (Ecosystem Dynamics and Sustainability), Institut Agro, Ifremer, Inrae, Rennes, France
5
Fisheries Commission, Accra, Ghana
* Corresponding authors: This email address is being protected from spambots. You need JavaScript enabled to view it.
; This email address is being protected from spambots. You need JavaScript enabled to view it.
Received:
25
August
2023
Accepted:
18
April
2024
Abstract
Habitats of three marine fish species of major interest (Epinephelus aeneus, Pseudotolithus senegalensis, and Pagellus bellottii) at both juvenile and adult life stages were identified and mapped in the western part of the Gulf of Guinea. Habitat suitability models (HSMs) were designed to quantify species- and stage-specific fish densities from scientific survey data collected for the last 40 years according to two environmental descriptors (bathymetry, distance to river mouth) and accounting for temporal contrasts. Delta general linear models were selected to fit with the 0-inflated distribution of the fish density data. Despite their high residual deviance, both the prediction accuracy and robustness of these HSMs were satisfactory. HSMs showed a strong influence of bathymetry on stage-specific fish distribution and a lower and non systematic influence of proximity to river mouths. The spatial distribution of juveniles of E. aeneus and P. senegalensis evidenced their concentration in shallow coastal nurseries. The adults of P. senegalensis were also located in nearshore habitats under estuarine influence, whereas adults of E. aeneus were spread towards deeper waters. P. bellottii did not rely on coastal fringes at juvenile or adult stages. Finally, model outputs showed a decrease in abundance in recent decades for all species at both life stages. These HSMs and maps reveal the importance of the coastal fringe as a critical essential habitat for two of the three studied species and the usefulness of space-based management measures to maintain populations and ensure sustainable fishing exploitation.
Key words: Fish habitat model / demersal fish / Gulf of Guinea / Epinephelus aeneus / Pseudotolithus senegalensis / Pagellus bellottii
Handling Editor: François LE LOC'H
© C.L. Quenum et al., Published by EDP Sciences 2024
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
1 Introduction
Estuarine and coastal ecosystems (ECEs) shelter a wide biological diversity and provide many ecosystem services (Barbier et al., 2011). Many marine species of fisheries interest carry out at least a part of their life cycle in ECEs (Seitz et al., 2014); in particular, the juvenile stage is frequently concentrated in these areas because they provide optimal growth conditions and protection from predators (Beck et al., 2001). This tendency was verified under temperate but also tropical latitudes for various ECEs, namely, mangroves (Manson et al., 2005), seagrasses (McDevitt-Irwin et al., 2016), and saltmarshes (Jänes et al., 2020), and the effects of these ECEs were found to be even stronger due to the combination of these complementary habitats and their connectivity (Barbier et al., 2011; Nagelkerken et al., 2015; Whitfield, 2017). For species relying on ECEs at the juvenile stage, both the size and the suitability of nursery habitats contribute to population renewal and productivity, and to sustain fisheries on adult stage in open ocean, at dramatically wider scale. However, ECEs usually faces a large panel of anthropogenic pressures (Halpern et al., 2012), especially in tropical zones (Friess, 2016). The cumulative effects of surface reduction and alteration of the quality of ECEs impact the suitability of juvenile habitats for many dependent exploited species (Brown et al., 2018; Worm et al., 2006). The management of exploited fisheries resources requires an ecological approach that considers not only fishing mortality but also recognizes the main role played by critical essential habitats (CEHs) and the related potential bottlenecks throughout the life cycle (Beck et al., 2001; Dahlke et al., 2020). The identification of nursery areas is a seminal crucial phase for the management of fisheries resources; it is essential for moving forward with the protection of these CEHs (Curnick et al., 2019).
The Gulf of Guinea is a biogeographical province with productive ecosystems, enhanced by large freshwater inflows and seasonal upwellings. These ecosystems sustain a large diversity of exploited marine species and related fisheries of main interest for local food supply (Hicks et al., 2019). The coastline of the Gulf of Guinea hosts a mosaic of ECEs (e.g., mud and sand flats, mangroves, saltmarshes) in river mouths, lagoons, deltas, bays or along the open coast (Okyere and Blay, 2020). Many exploited species in the Gulf of Guinea rely on these ECEs for their life cycle (John and Lawson, 1990). In particular, juveniles of many exploited marine species have been observed in and close to coastal lagoons and estuaries (Aheto et al., 2014; Bakari et al., 2016; Eugenia et al., 2019; Nunoo et al., 2006; Okyere et al., 2011; Okyere and Blay, 2020). Thus, the ECEs of the Gulf of Guinea contain nursery habitats (Cosme De Esteban et al., 2023), on which adverse effects of anthropogenic pressures (i.e., drop in area size due to land reclamation, degradation of habitat and water quality linked to eutrophication and pollution; cf. 2.1.) have been evidenced. However, the spatial extent of both juvenile and adult fish distributions, which is needed to identify CEHs, was still not delineated for exploited species in the Gulf of Guinea.
The present study focused on CEHs of three exploited species (E. aeneus, P. senegalensis, P. bellottii), selected regarding their high commercial value and landing volumes. The white grouper (Epinephelus aeneus, Saint-Hilaire, 1817) is the most important grouper (the common name “grouper” is usually given to fish in one of two large genera: Epinephelus and Mycteroperca) species in terms of catches. It has a very high local commercial and cultural value (this led to the inclusion of its drawing on a currency used banknote in West Africa). The white grouper landings, however, remain low: 127 tons in 2019; i.e. 0.1% of the demersal catches contributing to 0.4% of the commercial value in the study area (ZEE of Ivory Coast and Ghana, Sea Around Us data, Derrick 2020; Polido et al., 2020), and there is no information on its exploitation level. The cassava croaker (Pseudotolithus senegalensis, Valenciennes, 1833) is an economically important species (Sossoukpe et al., 2013b) with 7,432 tons of landings (calculated from Pseudotolithus spp. landings based on the proportion of P. senegalensis/Pseudotolithus spp. abundance in scientific survey data described in 2.2.); it represented 3.4% of the demersal catches in 2019 and 3.8% of the commercial values (Derrick, 2020; Polido et al., 2020). The cassava croaker is considered overexploited in the study area (FAO, 2022; Okyere and Blay, 2020). Red pandora (Pagellus bellottii, Steindachner, 1882) is the most widespread species in the Sparidae family; it represented 2.5% of the demersal catches (5,438 tons) and reached 4.6% of commercial value in 2019 (Derrick, 2020; Polido et al., 2020); it is considered at the full exploitation level in the study area (FAO, 2022; Kouame et al., 2020; Sylla et al., 2017).
White grouper and Cassava croaker are both fish species found in open lagoons and bays on the continental shelf of Gulf of Guinea (Bakari et al., 2016; Eugenia et al., 2019; Nunoo et al., 2006; Okyere and Blay 2020; Sossoukpe et al., 2013b). Red pandora are found in open waters (Kouame et al., 2020; Sylla et al., 2017).
The aim of this study was to map the spatial distribution of these three exploited species at the juvenile and adult life stages on the continental shelf of the western Gulf of Guinea. This involved (i) collating and standardizing scientific demersal trawl survey data from the last 4 decades in the study area, (ii) selecting habitat suitability models (HSMs) adapted to 0-inflated data distribution of species-specific fish abundance, and (iii) mapping these distributions with relation to environmental descriptors to identify CEHs for these three species.
2 Material and methods
2.1 Study site, selected species, fisheries and management
The coastal shelf of Côte d’Ivoire and Ghana spans along the western shore of the Gulf of Guinea (Fig. 1). From the shore to the 150 m depth (beginning of the continental slope), the continental shelf extent reaches 34,000 km2. This continental shelf is enriched by coastal upwellings during the cold marine season from June to October and January to February and by terrigenous inputs from rivers during the continental rain seasons, from June to July and then September to November. On this shelf, the fish community is highly diverse, with species of major interest for fisheries and food supply (Le Loeuff, 2001). The land‒sea interface hosts a mosaic of habitats: open and closed lagoons (Bakari et al., 2016; Laleye et al., 2007), deltas and estuaries (Okyere et al., 2011), and mangroves (Aheto et al., 2014; Osemwegie et al., 2016). These ECEs face cumulative anthropogenic pressures: mangrove surface loss (Abé and Affian, 1993; Osemwegie et al., 2016), eutrophication (Adingra and Kouassi, 2011), siltation and deoxygenation (Okyere and Nortey, 2018), and xenobiotic loadings (Koffi et al., 2014).
The studied species, i.e., White grouper, Cassava croaker and Red pandora are demersal fishes (Serranidae, Sciaenidae and Sparidae, respectively). Their distribution spans along the West African coasts to Angola, with some presences in the Mediterranean Sea for white grouper and red pandora (Kouame et al., 2020).
The cassava croaker is a gonochoric species; i.e., it maintains only one sex throughout its life cycle (male or female). White grouper and red pandora are hermaphroditic protogynous; i.e., they are born female and later become male. Sizes at first maturity (i.e., female maturity for protogynous species) vary among the three species (Tab. 1, see Supp. Tab. 1 for the related bibliographic review).
The management measures for the fishery resources in the study area are not mono specific. They focus on biological rest periods and consist of a temporary ban lasting 1 (July) to 2 (July–August) months for artisanal and industrial fishing fleets, respectively. The existence of a marine protected area for turtles also contributes to partial spatial conservation of marine fish populations.
 |
Fig. 1 Map of the study area in the Gulf of Guinea with river and lagoon mouths. 1: Cavally River; 2: San Pedro River; 3: Sassandra River; 4: Fresco Lagoon; 5: Grand-Lahou Lagoon; 6: Ebrié Lagoon; 7: Aby Lagoon; 8: Ankobra River; 9: Pra River; 10: Ayensu River; 11: Densu Delta; 12: Volta River. In the lower right corner: general location. |
Maturity information for the three studied species used to split juveniles and adults in the catches.
2.2 Trawl survey data
Both national and international scientific demersal trawl surveys carried out from 1978 to 2019 on the continental shelf of the study area were collected (temporal distribution, sampling effort, and technical characteristics of the fishing gear are given in Tab. 2). These surveys spanned from the coastal area (10 m depth) to the continental slope (Supp Fig. 1). Trawling speed and hauls duration were between [3–3.2] knots and [20–60] mins, respectively.
For each trawl haul (sample unit), the following information was collated: date, geographical position, area swept, and bathymetry. For the three studied species, catches were systematically identified, counted and weighted, but length structure was measured on a fraction of the trawl hauls only (on almost half of the trawl hauls where the species is caught for cassava croaker and red pandora and on less than 20% for white grouper; Tab. 3).
Demersal trawl survey data.
Proportion of fish caught, measured vs unmeasured.
2.3 Descriptors of spatiotemporal patterns in fish density used in HSMs
2.3.1 Environmental descriptors of fish habitat
Environmental data concern the descriptors used to model the spatial distribution for the three studied species at juvenile and adult stages; in this study, the data used are bathymetry and the proximity to habitats under riverine influence (Olds et al., 2012). See the Discussion for potential inclusion or other environmental descriptors.
2.3.1.1 Bathymetry
The bathymetry map (in m) is a raster with a spatial resolution of 15 arc seconds (NOAA, 2022; Supp. Fig. 2). Preliminary analysis showed that bathymetric distribution of the three species at juvenile or adult life stage was not monotonous (for an illustration, see the Results (Sect. 3.2.1) and the dome-shaped bathymetric distribution of white grouper and red pandora; Fig. 5). Regarding the amount of available trawl data (Tab. 2), this prevents the use of bathymetry as a covariate in habitat suitability models (HSMs) without overparametrization (Le Pape et al., 2014). Indeed, a choice was made to transform bathymetry into a class factor in HSMs. Bathymetry was split into ten categories (in m): [10–20], [20–30], [30–40], [40–50], [50–60], [60–70], [70–80], [80–90], [90, to 100] and >100, with a sufficient number of trawl hauls (Fig. 2) per class.
 |
Fig. 2 Number of samples (trawl hauls) per modality of descriptor. A: Bathymetry descriptor; B: Period descriptor; C: Proximity to river mouth descriptor. |
2.3.1.2 Proximity of river mouth or lagoon entrance
Twelve sites were selected as major estuarine entries in the study zone (Fig. 1). To describe the proximity with these connections to freshwater loadings, we used the shortest distance between the trawling stations and the river mouths. A threshold of distance was retained in two class factors: close to the river mouth (<50 km) and far from the river mouth (Fig. 2; Supp. Fig. 3). 50 km is a compromise between the number of hauls in each category and the effect detected on fish density in preliminary analyses. Preliminary analysis showed that in a range of distances from 30 to 60 km, fish density at the life stage was not significantly sensitive to this threshold. Note that regarding the narrowness of the continental shelf, the distance to the river mouth is not correlated with bathymetry (Supp. Fig. 4).
 |
Fig. 3 Mean weight by bathymetry class (in m) for measured vs unmeasured fish. |
 |
Fig. 4 Proportion of presence for real vs predicted values by bathymetry levels (in m). Illustration for juvenile. |
 |
Fig. 5 Prediction and mean effects of habitat descriptors. a:[10,20] m, b: [20,30] m, c: [30,40] m, d: [40,50] m, e: [50,60] m, f: [60,70] m, g: [70,80] m, h: [80,90] m, i: [90,100] m, j: >100m (except for cassava croaker for which f: [60,80] m and g: > 80m). |
2.3.2 Temporal stratification
Survey data spread over 41 discontinuous years. To incorporate the evolution of resources over time, the dataset was divided into two periods: one before 2000 and the other from 2000, i.e., 11 years of distinct trawl surveys (65.4% of the trawl hauls) and 8 years of distinct trawl surveys (35.6% of the trawl hauls), respectively (Fig. 2). The 2000 year was chosen as a good balance in terms of years and trawl hauls in the two categories.
2.4 Estimating the respective proportions of juveniles and adults in catches
For each species, catches needed to be split into juveniles and adults before modelling stage-specific distribution. When fish were measured, the proportion of juveniles regarding size at 1st maturity (Tab. 1) was directly derived from the length structure in catches. However, this was limited to a fraction of the samples (Tab. 3); for a large proportion of trawl hauls, only the total number of individuals caught and the total weight were available by species. The following method was used to categorize catches as juveniles or adults when length data were missing:
When a single individual was caught in a trawl haul for a studied species, its weight and the weight at 1st maturity were used to categorize it as juvenile or adult. The weight at 1st maturity (Tab. 1) was calculated regarding the size at maturity and the length and weight data available in the trawl survey dataset (Tab. 2).
When more than one individual was caught, a statistical approach was used. A preliminary analysis tested for potential biases in mean weight between measured/unmeasured fish data for the studied species. Then, for each species, generalized linear models (GLMs, McCullagh and Nelder, 1989) with Bernoulli distributions were used to estimate the species-specific proportion of juveniles in trawl hauls for which length structures are available (Eq. (1)):
 (1)
(1)
where J is the response variable, the proportion of juveniles in the sampling unit (trawl haul), X is the mean weight, ϵ is the residuals, and C is the number of fish measured.
The outputs of the 3 species-specific models were used to predict the proportion of juveniles in trawl hauls for which only mean weights are available (and catches are >1 fish) and infer the respective number of juveniles and adults in catches (Eq. (2)).
 (2)
(2)
where N is the number of juvenile fish in catches in the trawl haul, Ĵ is the predicted proportion of juveniles, and C is the number of fish caught in the trawl haul. A threshold is used when N < 1, N is replaced by 0 (absence). Number of adults = C–N.
In the whole dataset, catches by species were allocated in juveniles or adults; a complete dataset was available (2,077 trawl hauls), with:
The respective catch per unit of effort (relative density of fish in number per trawled surface) of juveniles and adults per species.
The environmental (i.e., bathymetry and proximity to freshwater mouth, derived from crossing the respective spatial maps Supp. Figs. 2 and 3 with the average geographic position of each trawl haul), and temporal (period) descriptors.
2.5 HSMs
Quantitative analysis consists of modelling nursery habitat suitability based on GLM methods (Le Pape et al., 2014; McCullagh and Nelder, 1989). The proportions of occurrence (Tab. 3) of white grouper, cassava croaker, and red pandora at all trawled stations are 33.26%, 21.76%, and 66.58%, respectively, and even less when the data are split by life stage. With regard to this 0-inflated distribution of catch data by species and life stage (Supp. Tab. 2), a delta model was used. A delta model is a combination of two submodels: one based on presence–absence and the other on abundance for non null densities (Le Pape et al., 2014; Trimoreau et al., 2013). Building this model needed three steps:
2.5.1 Submodel 1: probability of presence 
The presence–absence model aims to explain the probability of the presence of the fish for a given life stage, with a binomial distribution (Eq. (3)) based on presence/absence data (Boolean values: 0 when the relative density of fish in number per trawled surface is null, 1 otherwise).
 (3)
(3)
with Y0/1, the response variable, the probability of presence, βperiod, cofactor 1, period (2 categories; cf. 2.3.2.), βBathymetry, cofactor 2, bathymetry (in 10 categories; cf. 2.3.1.), β prox. to river mouth, cofactor 3, proximity to river mouth (2 categories; cf. 2.3.1.), ϵ0/1, residuals.
2.5.2 Submodel 2: positive densities Y+
The second submodel (Eq.( 4)) uses a Gaussian GLM on log-transformed positive densities to normalize the skewed distribution of the proxies of positive densities (CPUE). Preliminary analyses showed that this option was the best to describe the distribution of positive values in the dataset, as usually found for positive fish densities from survey data (Trimoreau et al., 2013).
 (4)
(4)
with log(Y+), log-transformed positive density in number per km2, β, explaining cofactors (cf. Eq. (3)), ε+, residuals.
2.5.3 Selection of explanatory variables
For the 2 submodels, for each species and life stage, the selection of explanatory factors was based on the Akaike information criterion (AIC), with a difference of 3 considered significant (Bozdogan, 1987). The submodels are compared one by one by adding an additional factor.
2.5.4 Coupling of both submodels: The delta model
Once the best submodels were selected for each species and life stage, they were coupled (Eq. (5)) by applying a correction to obtain an unbiased estimate from a linear model based on log-transformation (Trimoreau et al., 2013).
 (5)
(5)
with YS estimation of the density from the delta model, YS0/1, estimation of the probability of presence (Submodel 1, Eq. (3)), YS+, estimation of the positive log-density (Submodel 2, Eq. (4)),  , standard error of estimation of the log-transformed density (from Submodel 2).
, standard error of estimation of the log-transformed density (from Submodel 2).
2.5.5 Validation
To assess the goodness of fit (GOF) of the presence–absence models based on Boolean values (Submodel 1), the area under the curve (AUC) of the receiver operating characteristic (ROC) was used. The AUC is a threshold-independent measure aiming to assess the model performances at discriminating between presences and absences using a range of threshold values (Hanley and McNeil, 1982). The coefficient of determination (r2) was used to assess the GOF of Submodel 2 on positive densities.
As models with poor and/or biased prediction power could lead to spurious conclusions, their validation requires attention. To validate the models, their GOF was evaluated by comparing observed versus predicted values with a calibration/validation procedure. Each dataset (12, 2 submodels × 2 life stages × 3 species) was randomly split into two parts: calibration, 75% of the data, and validation, 25% of the remaining data. This random split was performed with respect to the relative amount of data within the different classes of factors (i.e., bathymetric classes, proximity to river mouth and period). For each species and life stage, binomial and positive models were fitted using the calibration dataset, and the GOF was tested both on the calibration and the validation datasets by comparing the model predictions with the data (predictions are the model-fitted values for the calibration dataset and the model estimates for the validation dataset; Trimoreau et al., 2013). This procedure was replicated 100 times, and the average calibration versus validation estimates were compared. Note that to overcome potential spatial correlation, this procedure was also tested after splitting the dataset into 2 geographical units (separated by economic exclusive zones, i.e. Côte d’Ivoire/Ghana). Similar results led us to consider the “simple” version without this spatial structure in the presented results.
2.6 Prediction with confidence intervals and mapping
In a delta model, the combined effects of the descriptors on the binomial distribution and on the positive log-data cannot be directly interpreted together (Le Pape et al., 2014). Combined mean effects have been computed to obtain relative effects of each modality of factor, with their associated uncertainty. Using a Monte–Carlo method (Jia, 2018), we proceeded in 3 steps: (i) generate 5000 random predictions from the presence–absence submodel  and the positive density submodel
and the positive density submodel 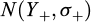 for each combination of modalities of the descriptors, (ii) couple the values obtained from the two submodels to obtain predicted densities (Eq. (6)), (iii) calculate the 5%, 50% and 95% quantiles of the distribution, i.e., the median and the associated 90% confidence interval, and show outputs by modality of the descriptor.
for each combination of modalities of the descriptors, (ii) couple the values obtained from the two submodels to obtain predicted densities (Eq. (6)), (iii) calculate the 5%, 50% and 95% quantiles of the distribution, i.e., the median and the associated 90% confidence interval, and show outputs by modality of the descriptor.
 (6)
(6)
Note that the estimation of the average positive density in equation (5) requires a correction used when logtransformation is applied  , Trimoreau et al., 2013). In equation (6), random predictions of positive densities are estimated by e (log(YS+)), without correction. Finally, quantile 50 of YS obtained here with equation (6) on 5000 random predictions is equivalent to YS estimated from equation (5).
, Trimoreau et al., 2013). In equation (6), random predictions of positive densities are estimated by e (log(YS+)), without correction. Finally, quantile 50 of YS obtained here with equation (6) on 5000 random predictions is equivalent to YS estimated from equation (5).
Habitat mapping consisted of combining the predicted densities (Eq. (5)) for each species and life stage with the geographic data (on each point of the spatial map, and for the two periods, the prediction of the density is done regarding the value of the spatial descriptors, bathymetry and proximity to the river mouths).
3 Results
3.1 Allocation of catches in life stage when length structure is lacking
The explained deviance of the extrapolation models of the proportion of juveniles in survey data reached 73%, 76%, and 62% for white grouper, cassava croaker, and red pandora, respectively (Tab. 4). Without different patterns in mean weight between measured and unmeasured data (Fig. 3), the models provide unbiased predictions of the proportion of juvenile fish in survey catches (Fig. 4). These models were used to predict the proportion of juveniles and complement the data with the number of juveniles and adults in catches when the length structure was not available.
Results of extrapolation models to estimate the proportion of juveniles.
3.2 Model selection, predictions and uncertainty
3.2.1 Selection of descriptors and model validation
The 3 descriptors, i.e., bathymetry, distance to river mouth, and period, were retained at least partly in the delta models for the three species (Tab. 5). For the three species and the two life stages, the two submodels very partly explained the deviance of the presence [10–28]% (min/max on the three species) and of the positive densities [3–18]%, respectively, with a better explanatory power for cassava croaker and worse for white grouper (Tab. 5).
The results of the juvenile and adult AUC data range from 0.701 ± 0.020 to 0.844 ± 0.015 except for the adult presence–absence submodel of white grouper (Tab. 6). The calibration and validation values are very close. The presence–absence submodels are therefore considered robust, except for white grouper adults. For the positive density submodels, the r2 coefficient is analytically equivalent to the percentage of explained deviance; thus, the performance of the models is similar for the calibration dataset (Tab. 6) than in the overall model fit (Tab. 5). Except for the adult white grouper, the degradation of r2 from calibration to validation data was below 20% for the three species and was considered reliable.
Habitat suitability models fit.
Calibration and validation results of habitat suitability sub-models (± standard deviation).
3.2.2 Model prediction and mapping
Overall, bathymetry had an important influence on habitat suitability (Tab. 5). The density of white grouper peaks in shallow areas for juveniles, is abundant between 10 and 50 m, whereas it is deeper for adults (Figs. 5 and 6). The cassava croaker is concentrated in the shallowest areas, below 20 m (Figs. 5 and 7). The red pandora has a wider distribution towards the open ocean, with a maximum abundance from 40 to 70 m and low density in the shallow coastal fringe (Figs. 5 and 8). For both cassava croaker and red pandora, the bathymetric distributions of juveniles and adults overlap (Fig. 5).
Distance to river mouth had an additional partial and moderate life-stage- and species-specific influence. Adults of white grouper are more abundant, and both life stages of cassava croaker more frequently occur close to the river mouths (Tab. 5; Fig. 5).
The period was significant, except for the presence of juveniles of white grouper and positive density for red pandora, indicating a general decrease in the abundance of both juvenile and adult fish for the three species (Tab. 5; Fig. 5).
 |
Fig. 6 Habitat maps of white grouper (E. aeneus), relative abundance (number of fish caught/km2). Regarding the low model accuracy for the adult stage, the adults map has to be considered as indicative. |
 |
Fig. 7 Habitat map of cassava croaker (P. senegalensis), relative abundance (number of fish caught/km2). |
 |
Fig. 8 Habitat map of red pandora (P. bellottii), relative abundance (number of fish caught/km2). |
4 Discussion
The present study aimed to model and map the spatial distribution of both juvenile and adult distributions of three fish species of main fishing interest in the western Gulf of Guinea. Data from trawl surveys carried out in the study area since the late 1970s were collated and pretreated to estimate the respective species-specific CPUE of juveniles and adults. The HSMs were developed to describe these CPUE with relation to environmental factors. Survey data (species- and life-stage-specific CPUE) were 0-inflated and prevented the use of conventional linear models. Delta GLMs combining two submodels for the probability of presence and positive densities, respectively, were applied to fit these 0-inflated data (Le Pape et al., 2014). These HSMs based on abiotic descriptors of the environment (bathymetry) and seascape (proximity to river mouths) and accounting for 2 time periods partly explained the spatial distribution of juveniles and adults and the temporal patterns in densities for the three species, with a large remaining part of the variability in distribution not explained by the descriptors.
4.1 Model limits and potential improvements
A poor level of explanatory power is common in marine fish HSMs, and residual variability is important (Le Pape et al., 2014). This was verified from the present case study: Uncertainties arise from observation processes (data paucity, small spatial scale and short-term temporal variability) but also from the lack of structuring environmental descriptors accounted for in model formulation.
The observation process is especially critical when the dataset merges various scientific surveys operated with various trawls and spanning 4 decades. Contrasts in trawl catchability and selectivity (Tab. 2) could have reduced the accuracy of CPUE estimates, especially for young juveniles. However, no reliable intercalibration information was available from these surveys carried out in various areas at different periods. In addition, the use of a single 1st size at maturity during the whole period (Tab. 1) could be a weakness. Accordingly, considering all individuals that were past their first maturity as adults led to considering protogynous fish as mature before male maturity and provided only a proxy of life-stage structure.
Accounting for a restricted number of environmental descriptors is another limit to the level of explanation of marine fish HSMs:
Sediment structure. Maps of sediment structures were not available with sufficient precision for the Ghanaian waters, and this parameter was not used as a habitat descriptor. However, HSMs based on a restricted dataset on Ivorian waters (Supp. Fig. 5) did not evidence a significant influence of this descriptor.
Additional longitudinal patterns. In response to differences in fisheries management between Ghana and Côte d’Ivoire, or to other underlying processes, species- and life stage-specific distributions of fish could present longitudinal patterns in the study area. However, tests using a “country” effect did not show any significant effect.
Temporal variability and hydrologic parameters. Salinity and/or nutrient concentration are potential complementary habitat descriptors, with, for instance, a better accuracy than seascape distance to the river mouth to describe the influence of estuarine enrichment (Trimoreau et al., 2013). In the context of climate change, a more accurate description of temporal patterns, integrating a descriptor of temperature, could also have improved model accuracy. However, these descriptors were not accounted for in the present analysis. The paucity of data mainly prevented this use. Hydrologic maps of surface temperature and salinity are not available for the first years of the time series of trawl surveys, and no information is available for the bottom waters, where these three demersal species are living.
Moreover, adding hydrology would imply to consider some temporal models, i.e., to account for interannual variability (Trimoreau et al., 2013) and to integrate the seasonal response of fish distribution to temperature, which is far from constant all year long. The paucity of trawl survey data (Tab. 1) did not allow for the development of such complex models. However, tests using a “season” effect in the model did not show any significant effect. Accounting for hydrologic variability is of course a major research avenue that would need further consideration in the future.
This modelling approach could thus be considered raw, providing only a proxy of fish density and partly explaining the average spatial distribution and temporal patterns in the long term, without integrating fine-scale distributions linked to biotic factors (food resources, Jänes et al., 2020; Le Pape et al., 2013) or multiscale temporal variability. However, an “out of the bag” cross-validation procedure (Le Pape et al., 2014) allowed us to validate HSMs robustness, except for adults of white grouper, for which model accuracy was lower. Even if more accurate datasets and environmental descriptors would certainly have refined the description and mapping, the present HSMs described some drivers of life-stage specific distribution for the three species and allowed us to map their distribution.
4.2 Species- and life stage-specific habitats and temporal patterns
The identification of habitats was carried out with two environmental descriptors: bathymetry and proximity to river mouths. Bathymetry is well known to structure the demersal fish distribution (Trimoreau et al., 2013) at both the juvenile and adult stages. The bathymetric factor was the most important driver of spatial distribution at both life stages for the three studied species. The influence of estuarine inputs is a complementary factor partly explaining the spatial distribution of marine species (Olds et al., 2012; Trimoreau et al., 2013), especially at juvenile stages (Beck et al., 2001). The enrichment of the coastal part of the continental shelf by river inputs from estuaries (Le Pape et al., 2013) makes the areas in their vicinity particularly suitable for the growth and survival of juveniles (Trimoreau et al., 2013). However, the contribution of this seascape component (Drakou et al., 2017; Nagelkerken et al., 2015) to the HSMs remained dramatically lower than that of bathymetry and logically concerned only the two coastal species.
Of the three studied species, white grouper was the one for which HSM performance was the lowest; it was robust for juveniles but less robust for adults. However, the similarity with conclusions drawn elsewhere for this species led us to validate HSM outputs and related maps. White grouper lives in ECEs such as open lagoons, bays, and estuaries (Bakari et al., 2016; Eugenia et al., 2019; Nunoo et al., 2006; Okyere and Blay 2020; Okyere and Nortey 2018; Sossoukpe et al., 2013b) at the juvenile stage. Out of the estuarine environment, juveniles are dramatically more abundant below 50 m of depth, confirming their high concentration on the coastal fringe of the continental shelf (Vally et al., 2023). At the adult stage, white grouper prefer deeper areas and have dramatically wider distributions; however, there is a preference for the proximity of estuaries (Vally et al., 2023).
Cassava croaker is frequently observed in ECEs (Nunoo et al., 2006; Okyere and Blay 2020; Okyere and Nortey 2018; Sossoukpe et al., 2013b). On the continental shelf, its spatial distribution is unimodal with a preference for shallow waters (<20 m) at both juvenile and adult stages and with a preference for the vicinity of estuaries (FAO, 1991). This species uses nearshore areas under riverine influence as nurseries, adult feeding and spawning areas (Sossoukpe et al., 2013a).
Red pandora live in the open sea (Kouame et al., 2020; Sylla et al., 2017). Juveniles and adults share the same living area, mainly from 30 to 80 m, with no dependency on coastal habitats.
For the three species, abundance decreased after 2000 at both the juvenile and adult stages. This decrease could be linked to a large panel of factors, including environmental changes and habitat degradation (Brown et al., 2018), evidenced in coastal habitats of the study area: land reclamation and habitat loss (Abé and Affian, 1993; Osemwegie et al., 2016), eutrophication (Adingra and Kouassi, 2011), siltation and deoxygenation (Okyere and Nortey, 2018), and xenobiotic loadings (Koffi et al., 2014). However, in the study area, global fishery landings have increased in recent decades, with overexploitation for Pseudotolithus spp. (Derrick, 2020; FAO, 2022; Polido et al., 2020) and, more precisely, for cassava croaker (Okyere and Blay, 2020; Tia et al., 2017), and full exploitation for red pandora (FAO 2022; Kouame et al., 2020; Sylla et al., 2017). In addition to the unestimated impact of climate change and habitat degradation, the impact of fisheries is partly responsive to the observed temporal changes in abundance for the three studied species.
4.3 Management recommendations
The quantitative identification and mapping of CEHs is a crucial preliminary step for an ecosystemic approach to preserve marine exploited resources (Le Pape et al., 2014). The protection of CEHs is important for the renewal of fish stock (Beck et al., 2001) and is needed in combination with fisheries management to sustain resources and fisheries (Worm et al., 2006).
The juveniles of white grouper and cassava croaker during their whole life cycle are concentrated at 0–50 m and 0–20 m, respectively. Thus, there is an urgent need to preserve/restore this coastal fringe of the continental shelf to enhance population renewal for these two species. The impacts of local anthropogenic disturbance on nearshore-dependent species are well documented (Brown et al., 2018). These disturbances, evidenced in the study area (cf. 4.2), could have detrimental impacts at the population level for production and exploitation. Local habitat degradation could impact populations of species residing in coastal habitats throughout their life cycle, such as cassava croaker, but could also have detrimental consequences at a dramatically larger scale for species concentrated in ECEs at the juvenile stage (Archambault et al., 2018; Drakou et al., 2017), such as white grouper. However, the consequences of habitat degradation for exploited fish population renewal remain poorly understood (Brown et al., 2018).
At the same time, there is a need to regulate fishing in the coastal part of the continental shelf. Pushing the fishing effort beyond CEHs using protected areas could be an effective option to protect nursery grounds for white grouper, with a positive impact on fish stocks without pushing fishermen out of fishing grounds for adults (Mesnildrey et al., 2013). However, this would imply restricting access to both juvenile and adult stages for cassava croaker. This would dramatically reduce its overexploitation level (FAO, 2022; Okyere and Blay, 2020) but will prevent sustaining fisheries landings. Regulation of the mesh sizes of the fishing gear targeting these species in the coastal fringe could be a complementary option to avoid catching juveniles, (i) concentrated there for white grouper, and (ii) as juveniles represent a high proportion of the fish landings for Pseudotolithus spp. (Sossoukpe et al., 2013b; Wehye et al., 2017).
Supplementary material
Supp Fig. 1. Geographic position of the trawl hauls in scientific surveys used in this study.
Supp Fig. 2. Bathymetry map of the continental shelf in the study area.
Supp Fig. 3. Buffer of 50 km from the entrance of river mouths to the continental shelf covering in the study area.
Supp Fig. 4. Number of trawl hauls per class of bathymetry x descriptor of proximity to the river mouths.
Supp Fig. 5. Sediment map of the continental shelf on the study zone.
Supp Table 1. Bibliographic review of the size at maturity of the three studied species.
Supp Table 2. Proportion of trawl hauls with the presence of the specie based on the total trawl hauls, including catches in life stages where the length structure was estimated.
Access Supplementary MaterialAcknowledgement
The authors are grateful to the Centre de Recherches Océanologiques (Côte d’Ivoire) and the Department of Fishery (Ghana) for providing their data for this study and all who were contacted during data procurement. This work was funded by the European Project Demerstem (PESCAO and ECOWAS). The authors thank Florian Quemper for his help regarding stock exploitation status of the studied species and Benedicta Tigyimah for proofreading the English language. We are also grateful to the editor and the three anonymous reviews who significantly helped us to improve the quality of this manuscript.
Data availability statement
The authors do not have permission to share data.
References
- Abé J, Affian K. 1993. Morphology and tourist infrastructures of the Ivorian Coast BT, in: P.P. Wong (Ed.), Tourism vs Environment: The Case for Coastal Areas. Dordrecht: Springer Netherlands, pp. 99–108. [Google Scholar]
- Adingra AA, Kouassi AM. 2011. Pollution en lagune Ebrié et ses impacts sur l’environnement et les populations riveraines. Fiches Tech Doc Vulg 53: 48–53. [Google Scholar]
- Aheto DW, Okyere I, Asare NK, Dzakpasu MFA, Wemegah Y, Tawiah P, Dotsey-Brown J, Longdon-Sagoe M. 2014. A survey of the benthic macrofauna and fish species assemblages in a mangrove habitat in Ghana. West Afr J Appl Ecol 22: 1–15. [Google Scholar]
- Archambault B, Rivot E, Savina M, Le Pape O. 2018. Using a spatially structured life cycle model to assess the influence of multiple stressors on an exploited coastal-nursery-dependent population. Estuar Coast Shelf Sci 201: 95–104. [CrossRef] [Google Scholar]
- Bakari C, Léonard T, Toussaint J-TG, Tidiani K, Kouamelan P-E. 2016. Fish assemblage structure in the Tropical Coastal Lagoon of Grand Lahou (Côte d’Ivoire, West Africa). Int J Fish Aquac 8: 1–13. [CrossRef] [Google Scholar]
- Barbier EB, Hacker SD, Kennedy C, Koch EW, Stier AC, Silliman BR. 2011. The value of estuarine and coastal ecosystem services. Ecol Monogr 81: 169–193. [CrossRef] [Google Scholar]
- Beck MW, Heck KL, Able KW, Childers DL, Eggleston DB, Gillanders BM, Halpern B, Hays CG, Hoshino K, Minello TJ, Orth RJ, Sheridan PF, Weinstein MP. 2001. The identification, conservation, and management of Estuarine and Marine Nurseries for Fish and Invertebrates: a better understanding of the habitats that serve as nurseries for marine species and the factors that create site-specific variability in nurse. Bioscience 51: 633–641. [CrossRef] [Google Scholar]
- Bozdogan H. 1987. Model selection and Akaike’s Information Criterion (AIC): the general theory and its analytical extensions. Psychometrika 52: 345–370. [CrossRef] [MathSciNet] [Google Scholar]
- Brown EJ, Vasconcelos RP, Wennhage H, Bergström U, Støttrup JG, van de Wolfshaar K, Millisenda G, Colloca F, Le Pape O. 2018. Conflicts in the coastal zone: human impacts on commercially important fish species utilizing coastal habitat. ICES J Mar Sci 75: 1203–1213. [CrossRef] [Google Scholar]
- Cosme De Esteban M, Haroun R, Tuya F, Abreu AD, Otero-Ferrer F. 2023. Mapping marine habitats in the Gulf of Guinea: a contribution to the future establishment of Marine Protected Areas in Principe Island. Reg Stud Mar Sci 57: 102742. [Google Scholar]
- Curnick DJ, Pettorelli N, Amir AA, Balke T, Barbier EB, Crooks S, Dahdouh-Guebas F, Duncan C, Endsor C, Friess DA, Quarto A, Zimmer M, Lee SY. 2019. The value of small mangrove patches. Science (80-.) 363: 239–239. [CrossRef] [PubMed] [Google Scholar]
- Dahlke FT, Wohlrab S, Butzin M, Pörtner H-O. 2020. Thermal bottlenecks in the life cycle define climate vulnerability of fish. Science (80-.). 369: 65–70. [CrossRef] [PubMed] [Google Scholar]
- Derrick B. 2020. Côte d’Ivoire: Updated catch reconstruction for 2011–2018. Updat. to 2018 1950–2010 Mar. Catch Reconstr. Sea Around Us Part I − Africa, Antarct Eur North Atl 28: 31–34. [Google Scholar]
- Drakou EG, Pendleton L, Effron M, Ingram JC, Teneva L. 2017. When ecosystems and their services are not co-located: oceans and coasts. ICES J Mar Sci 74: 1531–1539. [CrossRef] [Google Scholar]
- Eugenia BB, Armah AK, Dankwa HR. 2019. Fish as bioindicators of habitat degradation in coastal lagoons of Ghana. Bonorowo Wetl. 9. [Google Scholar]
- FAO. 1991. The “GUINEA 90” survey. CECAF|ECAF, Fishery Committee for the Eastern Central Atlantic. [Google Scholar]
- FAO. 2022. Fishery Committee for the Eastern Central Atlantic (CECAF), Scientific Sub-Committee, Ninth Session, Demersal resources Working Group South 2022-meeting summary. Nouakchott, Mauritania. [Google Scholar]
- Friess DA. 2016. Mangrove forests. Curr Biol 26: R746– R748. [CrossRef] [PubMed] [Google Scholar]
- Halpern BS, Longo C, Hardy D, McLeod KL, Samhouri JF, Katona SK, Kleisner K, Lester SE, O’Leary J, Ranelletti M, Rosenberg AA, Scarborough C, Selig ER, Best BD, Brumbaugh DR, Chapin FS, Crowder LB, Daly KL, Doney SC, Elfes C, Fogarty MJ, Gaines SD, Jacobsen KI, Karrer LB, Leslie HM, Neeley E, Pauly D, Polasky S, Ris B, St Martin K, Stone GS, Sumaila UR, Zeller D. 2012. An index to assess the health and benefits of the global ocean. Nature 488: 615–620. [CrossRef] [PubMed] [Google Scholar]
- Hanley JA, McNeil BJ. 1982. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 143: 29–36. [Google Scholar]
- Hicks CC, Cohen PJ, Graham NAJ, Nash KL, Allison EH, D’Lima C, Mills DJ, Roscher M, Thilsted SH, Thorne-Lyman AL, MacNeil MA. 2019. Harnessing global fisheries to tackle micronutrient deficiencies. Nature 574: 95–98. [CrossRef] [PubMed] [Google Scholar]
- Jänes H, Macreadie PI, Nicholson E, Ierodiaconou D, Reeves S, Taylor MD, Carnell PE. 2020. Stable isotopes infer the value of Australia’s coastal vegetated ecosystems from fisheries. Fish Fish 21: 80–90. [CrossRef] [Google Scholar]
- Jia B. 2018. The application of Monte Carlo methods for learning generalized linear model. Biometrics Biostat Int J 7. [Google Scholar]
- John DM, Lawson GW. 1990. A review of mangrove and coastal ecosystems in West Africa and their possible relationships. Estuar Coast Shelf Sci 31: 505–518. [CrossRef] [Google Scholar]
- Koffi YB, Ahoussi KE, Kouassi AM, Biemi J. 2014. Ressources minières, pétrolières et gazières de la Côte d’Ivoire et problématique de la pollution des ressources en eau et des inondations. Geo Eco Trop 38: 119–136. [Google Scholar]
- Kouame AC, Sylla S, Yao SS, Tia CB, Loukou GA. 2020. Stock assessment of red Pandora Pagellus Bellottii (Steindachner, 1882) in the Ivorian continental shelf (West Africa). Agron. Africaine 32: 423–438. [Google Scholar]
- Laleye PA, Villanueva CM, Enstua-Mensah M, Moreau J. 2007. A review of the aquatic living resources in Gulf of Guinea lagoons, with particular emphasis on fisheries management issues. J Afrotropical Zool 123–136. [Google Scholar]
- Le Loeuff P. 2001. L’écosystème benthique au large de Grand-Bassam (Côte-d’Ivoire). Considérations sur les variabilités saisonnière, interannuelle et à long terme. Doc Sci du Cent IRD Bretagne 87: 27. [Google Scholar]
- Le Pape O, Delavenne J, Vaz S. 2014. Quantitative mapping of fish habitat: A useful tool to design spatialised management measures and marine protected area with fishery objectives. Ocean Coast Manag 87: 8–19. [CrossRef] [Google Scholar]
- Le Pape O, Modéran J, Beaunée G, Riera P, Nicolas D, Savoye N, Harmelin-Vivien M, Darnaude AM, Brind’Amour A, Le Bris H, Cabral H, Vinagre C, Pasquaud S, França S, Kostecki C. 2013. Sources of organic matter for flatfish juveniles in coastal and estuarine nursery grounds: a meta-analysis for the common sole (Solea solea) in contrasted systems of Western Europe. J Sea Res 75: 85–95. [CrossRef] [Google Scholar]
- Manson FJ, Loneragan NR, Skilleter GA, Phinn SR. 2005. An evaluation of the evidence for linkages between mangroves and fisheries: a synthesis of the literature and identification of research directions, in: Oceanography and Marine Biology. CRC Press, pp. 493-524. [Google Scholar]
- McCullagh P, Nelder JA. 1989. Generalized Linear Models, 2nd edn. London, New York, Monographs on Statistics and Applied Probability 37. [CrossRef] [Google Scholar]
- McDevitt-Irwin J, Iacarella J, Baum J. 2016. Reassessing the nursery role of seagrass habitats from temperate to tropical regions: a meta-analysis. Mar Ecol Prog Ser 557: 133–143. [CrossRef] [Google Scholar]
- Mesnildrey L, Gascuel D, Le Pape O. 2013. Integrating Marine Protected Areas in fisheries management systems: some criteria for ecological efficiency. Aquat Living Resour 26: 159–170. [CrossRef] [EDP Sciences] [Google Scholar]
- Nagelkerken I, Sheaves M, Baker R, Connolly RM. 2015. The seascape nursery: a novel spatial approach to identify and manage nurseries for coastal marine fauna. Fish Fish 16: 362–371. [CrossRef] [Google Scholar]
- NOAA. 2022. ETOPO 2022 15 Arc-Second Global Relief Model. Natl. Centers Environ. Inf. [Google Scholar]
- Nunoo F, Eggleston D, Vanderpuye C. 2006. Abundance, biomass and species composition of nearshore fish assemblages in Ghana, West Africa. Afr J Mar Sci 28: 689–696. [CrossRef] [Google Scholar]
- Okyere I, Aheto DW, Aggrey-fynn J. 2011. Comparative ecological assessment of biodiversity of fish communities in three coastal wetland systems in Ghana 1, 178–188. [Google Scholar]
- Okyere I, Blay J. 2020. Assessment of the fishery, growth and mortality characteristics of the cassava croaker, Pseudotolithus senegalensis (Sciaenidae) from coastal waters of Ghana. Reg Stud Mar Sci 39: 101425. [Google Scholar]
- Okyere I, Nortey D. 2018. Assessment of the Ecological Health Status of River Pra Estuary (Ghana) and Adjoining Wetland using Physico-chemical Conditions and Macroinvertebrate Bioindicators. West Afr J Appl Ecol 26: 44–55. [Google Scholar]
- Olds AD, Connolly RM, Pitt KA, Maxwell PS. 2012. Habitat connectivity improves reserve performance. Conserv Lett 5: 56–63. [CrossRef] [Google Scholar]
- Osemwegie I, Hyppolite DN, Stumpp C, Reichert B, Biemi J. 2016. Mangrove Forest Characterization in Southeast Côte d’Ivoire. Open J Ecol 06: 138–150. [CrossRef] [Google Scholar]
- Polido R, Noël S-L, Amador K. 2020. Ghana: Updated catch reconstruction for 2011–2018. In: Derrick B, Khalfallah M, Relano V, Zeller D, Pauly D. (Eds.), Fisheries Centre Research Report. pp. 40–45. [Google Scholar]
- Seitz RD, Wennhage H, Bergström U, Lipcius RN, Ysebaert T. 2014. Ecological value of coastal habitats for commercially and ecologically important species. ICES J Mar Sci 71: 648–665. [CrossRef] [Google Scholar]
- Sossoukpe E, Nunoo FKE, Adite A. 2013a. Population structure and reproductive parameters of the Cassava croaker, Pseudotholithus senegalensis (Pisces, Valenciennes, 1833) in nearshore waters of Benin (West Africa) and their implications for management. Int J Dev Res 3: 37–45. [Google Scholar]
- Sossoukpe E, Nunoo FKE, Ofori-Danson PK, Fiogbe ED, Dankwa HR. 2013b., Growth and mortality parameters of P. senegalensis and P. typus (Sciaenidae) in nearshore waters of Benin (West Africa) and their implications for management and conservation. Fish Res 137: 71–80. [CrossRef] [Google Scholar]
- Sylla S, Kouakou KF, Tia CB, Yao SS, Atse BC. 2017. The spatial distribution of coastal fish assemblage in Côte d’Ivoire’s Exclusive Economic Zone (EEZ), West Africa. J Appl Biosci 108: 10519. [CrossRef] [Google Scholar]
- Tia CB, Konan KJ, Sylla S, Kouamelan EP, Atse BC. 2017. Population parameters and stock assessment of the cassava croaker Pseudotolithus senegalensis (Valenciennes, 1833) in the Coastal Waters of Côte d’Ivoire. Int J Sci Res Methodol Hum J 6: 79–95. [Google Scholar]
- Trimoreau E, Archambault B, Brind’Amour A, Lepage M, Guitton J, Le Pape O. 2013. A quantitative estimate of the function of soft-bottom sheltered coastal areas as essential flatfish nursery habitat. Estuar Coast Shelf Sci 133: 193–205. [CrossRef] [Google Scholar]
- Vally Y. El, Quenum CL, Thiaw M, Guitton J, Meissa B, Le Pape O. 2023. Quantitative mapping of effective habitats for the White grouper Epinephelus aeneus (Geoffroy Saint-Hilaire, 1817) in North West Africa. Reg Stud Mar Sci 65: 103067. [Google Scholar]
- Wehye AS, Ofori Danson PK, Lamptey AM. 2017. Population dynamics of Pseudotolithus senegalensis and Pseudotolithus typus and their implications for management and conservation within the coastal waters of Liberia. Fish Aquac J 08. [Google Scholar]
- Whitfield AK. 2017. The role of seagrass meadows, mangrove forests, salt marshes and reed beds as nursery areas and food sources for fishes in estuaries. Rev Fish Biol Fish 27: 75–110. [CrossRef] [Google Scholar]
- Worm B, Barbier EB, Beaumont N, Duffy JE, Folke C, Halpern BS, Jackson J.BC, Lotze HK, Micheli F, Palumbi SR, Sala E, Selkoe KA, Stachowicz JJ, Watson R. 2006. Impacts of biodiversity loss on ocean ecosystem services. Science (80-.) 314: 787–790. [Google Scholar]
Cite this article as: Quenum CL, Vally YE, Tapé J, Guitton J, Ekuban EF, Kouakou ND, Le Pape O. 2024. Identification of critical essential habitat for demersal fish in the Gulf of Guinea. Aquat. Living Resour. 37: 7
All Tables
Maturity information for the three studied species used to split juveniles and adults in the catches.
Calibration and validation results of habitat suitability sub-models (± standard deviation).
All Figures
|
Fig. 1 Map of the study area in the Gulf of Guinea with river and lagoon mouths. 1: Cavally River; 2: San Pedro River; 3: Sassandra River; 4: Fresco Lagoon; 5: Grand-Lahou Lagoon; 6: Ebrié Lagoon; 7: Aby Lagoon; 8: Ankobra River; 9: Pra River; 10: Ayensu River; 11: Densu Delta; 12: Volta River. In the lower right corner: general location. |
| In the text | |
|
Fig. 2 Number of samples (trawl hauls) per modality of descriptor. A: Bathymetry descriptor; B: Period descriptor; C: Proximity to river mouth descriptor. |
| In the text | |
|
Fig. 3 Mean weight by bathymetry class (in m) for measured vs unmeasured fish. |
| In the text | |
|
Fig. 4 Proportion of presence for real vs predicted values by bathymetry levels (in m). Illustration for juvenile. |
| In the text | |
|
Fig. 5 Prediction and mean effects of habitat descriptors. a:[10,20] m, b: [20,30] m, c: [30,40] m, d: [40,50] m, e: [50,60] m, f: [60,70] m, g: [70,80] m, h: [80,90] m, i: [90,100] m, j: >100m (except for cassava croaker for which f: [60,80] m and g: > 80m). |
| In the text | |
|
Fig. 6 Habitat maps of white grouper (E. aeneus), relative abundance (number of fish caught/km2). Regarding the low model accuracy for the adult stage, the adults map has to be considered as indicative. |
| In the text | |
|
Fig. 7 Habitat map of cassava croaker (P. senegalensis), relative abundance (number of fish caught/km2). |
| In the text | |
|
Fig. 8 Habitat map of red pandora (P. bellottii), relative abundance (number of fish caught/km2). |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.