| Issue |
Aquat. Living Resour.
Volume 36, 2023
Topical Issue - NORA 4 - Native Oyster Restoration Alliance 4th Conference
|
|
|---|---|---|
| Article Number | 13 | |
| Number of page(s) | 18 | |
| DOI | https://doi.org/10.1051/alr/2023006 | |
| Published online | 08 May 2023 | |
Review Article
Fifty years of research to counter the decline of the European flat oyster (Ostrea edulis): a review of French achievements and prospects for the restoration of remaining beds and revival of aquaculture production
1
LEMAR, Ifremer, CNRS, IRD, UBO, 29840 Argenton en Landunvez, France
2
MARBEC, Univ Montpellier, CNRS, Ifremer, IRD, 34000 Montpellier, France
3
Ifremer, ASIM Adaptation et Santé des Invertébrés Marins, 17390 La Tremblade, France
4
Département Ressources Biologiques et Environnement, Ifremer, ZI de la pointe du diable, CS 10070, 29280 Plouzané, France
* Corresponding author: This email address is being protected from spambots. You need JavaScript enabled to view it.
Handling Editor: Pauline Kamermans
Received:
20
July
2022
Accepted:
13
March
2023
Abstract
In the fifty years since the introduction of the Pacific oyster Crassostrea gigas and the first reports of the parasites Marteilia refringens and Bonamia ostreae in European waters, numerous research projects dedicated to the native European flat oyster Ostrea edulis have been conducted, notably in France. Most of these projects have been dedicated to developing controlled reproduction and hatchery technology for seed production, examining pathological aspects to understand and control diseases, and using genetics to develop resistant lines. While the long-term objective of most studies has been to revive the aquaculture production of O. edulis, a smaller number have addressed the ecology of local remnant beds and reefs in France. This article provides an overview of the major outcomes of all these projects, focusing on results obtained in France and prospects for future work there, taking into account the rising interest in increasing aquaculture production and ecological motivation to restore declining populations as part of the framework of the Native Oyster Restoration Alliance (NORA) and in line with UN Decade for Ecosystem Restoration.
Key words: European flat oyster / genetics / pathology / ecology / restoration
© S. Pouvreau et al., Published by EDP Sciences 2023
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
1 Introduction
The flat oyster Ostrea edulis is the only native oyster species in Europe apart from some dwarf oysters present in Spain and Portugal (see Prado et al., 2022). The so-called ‘Portuguese oyster’, Crassostrea angulata, has instead been shown to be native to southern Asia (Boudry et al., 1998) and the Pacific oyster, Crassostrea gigas, was originally introduced from Japan (Grizel and Héral, 1991).
Among edible marine bivalves, O. edulis has long been known as a delicacy, leading to a decline in its stocks along European shores from overharvesting since the middle ages (Dupont and Rouzeau, 2015). Beds and reefs used to occupy large areas on the shore and in the subtidal zone on many coasts of France, especially in Brittany (Joubin, 1908), but they have almost disappeared after several centuries of over-exploitation and are now critically endangered (Duchene et al., 2015). This particular habitat was called Huîtrières in old French. Surprisingly, with the loss of this habitat, this term has also disappeared from the French language. The forgetting of this term over the generations is a symptom of a kind of collective amnesia, which was similarly reported by Alleway and Connell (2015) in their work on the history of the flat oyster O. angasi in Australia.
After the depletion of natural beds in France, oyster farming was developed from the end of the 19th century and yearly production quickly reached more than 20,000 tons in the first half of the 20th century. However, high mortality occurred from the 1970 s onwards with the emergence of two parasites, Marteilia refringens and Bonamia ostreae (see Sect. 3.1). As a result, French aquaculture production of O. edulis declined from up to 30,000 tons per year at the end of the 1960 s down to less than 2000 tons today. France is and has always been by far the main producer of flat oysters in Europe and the French government recently set the ambitious target of restoring French aquaculture production of O. edulis to 8,000 tons in 2030 (Anonyme, 2021).
Since these outbreaks, a large number of studies in France have been dedicated to better characterizing O. edulis diseases and to attempting to breed more resilient flat oyster strains (see Tab. 1). In a recent bibliographic analysis on O. edulis (Bertolini and Pastres, 2022) it is striking that Bonamia ostreae is the top listed keyword, with at least 13% of the studies gathered being about Bonamia infection. Surprisingly, there have been very few studies in France on the ecology and physiology of this species and its habitat since the 1970 s, as shown in Table 1. However, there has been a recent revival of interest in these topics driven by the wish to conduct ecological restoration of the species (zu Ermgassen et al., 2020). In addition, Colsoul et al. (2021) recently reviewed the current state of knowledge about the biological basis and technologies of O. edulis seed production. Most of this knowledge was gleaned from projects dedicated to aquaculture and now contributes to an increasing supply of seed for use in restoration projects.
In this context, the present article aims to review the major outcomes of research projects on O. edulis genetics and pathology conducted since the 1970 s. We especially examined research articles and reports available in Archimer, Ifremer's publicly available institutional repository (https://archimer.ifremer.fr/). Our review also provides a summary of the current state of the wild populations of flat oysters still present in France and the main causes of their decline in order to identify the most suitable restoration methods for French coastal environments under the European framework of the Native Oyster Restoration Alliance (NORA). zu Ermgassen et al. (2020) identified 40 top questions on oyster restoration in Europe, which they grouped into ten themes, including those we cover here: diversity and population differentiation, disease management, baselines and causes of decline, and restoration methods. In practical terms, the present article aims to provide information and guidelines to restore declining wild populations in France and to revive French aquaculture of this emblematic species.
Non-exhaustive inventory of research and development work (published articles and reports) carried out in France since the emergence of the epizootics linked to the parasites Marteilia refringens and Bonamia ostreae. Most cited references are available via https://archimer.ifremer.fr/ or on request from the French technical centres CAPENA, SMIDAP or SMEL.
2 Genetics
As early as the 1980 s, French research started to focus on genetic characterization of wild populations of flat oysters, thanks to the development of the first molecular markers (Tab. 1). In the 1990 s, the first attempts to perform selective breeding to overcome mortalities caused by Bonamiosis were successful. Then, in the 2000 s, DNA markers became available, allowing more in-depth investigations both in population genetics and selective breeding. The objectives were to obtain better descriptions of the resources available in the wild to be potentially used in restoration or breeding and to develop tools to assist these activities.
2.1 Wild populations
From a population genetics point of view, marine bivalve molluscs are characterized by very high fecundity in the gametic phase, strong potential for long-distance passive dispersal in the planktonic larval phase, and large population sizes in the benthic adult phase. As a result, a high genetic diversity is expected in populations, guaranteeing an adaptive potential in the face of constraints encountered in the wild. Significant exchanges between populations are also expected and, therefore, a weak differentiation between populations. However, many studies have revealed a more complex situation, as notably shown in O. edulis.
The structuration of O. edulis genetic diversity at the European level was first analysed using allozymes (Blanc et al., 1986; Johannesson et al., 1989; Jaziri, 1990; Saavedra et al., 1993, 1995), mitochondrial DNA (mtDNA) (Diaz-Almela et al., 2004) and microsatellites (Launey et al., 2002; Sobolewska and Beaumont 2005; Beaumont et al., 2006; Lallias et al., 2009a; Vera et al., 2016). All the studies generally agreed there was a weak but significant genetic differentiation between Atlantic and Mediterranean populations, following an isolation by distance model (Saavedra et al., 1995; Launey et al., 2002; Diaz-Almela et al., 2004). Hence, despite a supposedly high gene flow due to pelagic larval dispersal, O. edulis populations do not constitute a single panmictic unit (Launey et al., 2002), and studies using larger numbers of microsatellite markers (Vera et al., 2016) or SNP markers (Lapègue et al., 2014, Lapègue et al., 2023) identified several clusters in Europe. Two of these clusters are present in France, one along the English Channel and Atlantic coast, and the other along the Mediterranean coast. Gutierrez et al. (2017), using more than 2000 SNP markers, confirmed a high homogeneity of populations along the English Channel and Atlantic coast. The observed diversity is often higher in the Mediterranean Sea than along the Atlantic coasts (Saavedra et al., 1995, Launey et al., 2002; Diaz-Almela et al., 2004), which may result from a lower impact of human activities or biological stressors. However, the structure of Mediterranean Sea populations requires more thorough investigation.
Moreover, genomic islands of differentiation were detected, mainly in the form of two groups of linked markers, which could indicate the presence of chromosomal rearrangements in the O. edulis genome. These two genomic islands and the most differentiated loci also tended to show a parallel pattern of differentiation, that is to say comparable allelic frequencies between the North Sea populations and the Eastern Mediterranean and Black Sea populations, which was defined as a genetic parallelism (Lapègue et al., 2023). This genetic parallelism could be the sign of a shared evolutionary history of the two groups of populations despite them being at opposite edges of the present distribution of the species (Bierne et al., 2013).
At the level of the whole species range, understanding potential genetic parallelisms and more generally the genetic structure of wild and cultured populations is of prime interest to better understand their evolutionary history and to better manage stocks (Bernatchez et al., 2019). Hence, the identified mosaic of clusters is genuine and these clusters can be considered as Evolutionary Significant Units (ESUs) that need to be managed with dedicated conservation strategies. In addition, genetic information must be considered early on when selecting the geographic origin of oysters to be transferred or reproduced in hatcheries for subsequent restocking. Genetic monitoring protocols then need to be conducted in order to follow the evolution of the genetic diversity because this is generally considered to contribute to a species' ability to adapt to environmental changes (DeWoody et al., 2021).
2.2 Captive populations and selective breeding
The high fecundity of marine bivalves combined with the unpredictable and highly variable success of reproduction and recruitment (sweepstakes reproduction) they experience in the wild lead to a greater decoupling between Ne (effective population size) and N (census size) than in other organisms, which could explain why a higher segregating load can be maintained for a given silent mutation effective size (Harrang et al., 2013). Hence, the effective number of breeders can be low, especially in hatchery stocks (Lallias et al., 2010a) but also in wild conditions (Lallias et al., 2010b), supporting the hypothesis of sweepstake reproductive success in this species (Hedgecock et al., 2007; Taris et al., 2009; Hedgecock and Pudovkin, 2011). This low number of effective breeders was observed as soon as the first trials were made to develop breeding programs for resistance to bonamiosis (Launey et al., 2001). This trait has been the main one focused on by selective breeding programs because of the drastic decrease in production caused by the parasite Bonamia ostreae. Such programs were first developed in France and Ireland in the mid-1980 s (Baud et al., 1997; Culloty et al., 2001; Martin et al., 1992, Naciri-Graven et al., 1998, 1999), based on mass selection and parasite inoculation (Mialhe et al., 1988). In order to restore a sufficient level of genetic diversity, selective breeding in France was later reoriented to follow a within-family selection scheme (Bédier et al., 2001). Studies showed significant genetic gains for survival to bonamiosis but did not manage to provide heritability estimates. New tools make it possible to better characterize the genome of this species within the genus and the Ostreidae more generally (Leitao et al., 2004, Danic-Tchaleu et al., 2011), with efforts focused on the development of genetic markers and low-to-medium density genetic maps (Lallias et al., 2007; Lapègue et al., 2014) in the attempt to link markers to traits of interest. Hence, QTLs (Quantitative Trait Loci) associated with resistance to bonamiosis were identified and some were confirmed in different genetic backgrounds (Lallias et al., 2008, 2009b; Harrang et al., 2015), explaining up to 17% of the phenotypic variance of this trait. More recently, thanks to the development of a medium density Affimetrix Axiom array (Gutierrez et al., 2017) and the availability of transcriptome databases (Pardo et al., 2016; Ronza et al., 2018), notably revealing genes differentially expressed between naïve oysters and oysters of strains exposed to the parasite since the early 1980 s (including one exposed population in Brittany), 22 SNPs markers were identified as associated to the divergent selection between the two groups of wild populations (Vera et al., 2019). As these markers were located within a single genomic region, this suggests a major QTL associated with bonamiosis resistance under selection in wild populations exposed to bonamia (Sambade et al., 2022). Quantifying phenotypic and genetic variance associated with QTLs remains challenging, notably because it has been shown that the expression of some immune-related genes is variable over time (Morga et al., 2011) and, consequently, different QTLs might be identified at different developmental stages.
2.3 Perspectives for future genetics research
First, a better characterization needs to be made of the boundaries between the different genetic units in the European flat oyster. This is particularly true in the English Channel and on the Atlantic coasts of Spain and Portugal from which more samples are required. This is clearly even more challenging in the Mediterranean and Black Seas where many more coasts still need to be covered. Even once this information on genetic boundaries is known, the origin of oysters for restocking if an ESU became critically endangered would still need to be discussed for each case. A choice would then need to be made between introducing animals from (i) the genetically (and often geographically) closest populations as indicated by most of the genome, (ii) populations sharing alleles from specific loci indicating potential common adaptation and/or a shared ancient history, or (iii) an admix of both.
Second, efforts should be made to identify genes involved in the immune response of the flat oyster to the parasite (Morga et al., 2012: 2017), combined with the new genomic tools available such as whole genome sequence and chromosome level assembly (Bean et al., 2022; Boutet et al., 2022; Gundappa et al., 2022; Li et al., 2023). This should rapidly make it possible to precisely characterize the genetic basis of the resistance to bonamiosis and to consider the applications of this knowledge in breeding programs. Furthermore, as implemented in the Pacific oyster, research on epigenetic modifications and their potential transgenerational effects on traits of interest, such as resistance to disease, is also needed (Fallet et al., 2020).
While experimental selection clearly showed that improving survival of Bonamiosis is feasible and allows substantial gains, no national breeding programme has yet been launched for O. edulis in France, although some regional initiatives are being taken. However, thanks to the availability of genomic tools, and despite the difficulty of raising flat oysters, genomic selection is now a realistic perspective. However, it still remains to be implemented in the flat oyster industry so as to be of benefit to restoration programs.
3 Pathology
Diseases have long been recognized as being among the most important threats to marine molluscs, including the flat oyster O. edulis. Described as the agent of the ‘shell disease', the fungus Ostracoblabe implexa was initially reported in Arcachon in 1878 but then quickly spread to Brittany between 1947 and 1960 (Marteil, 1976). This fungus grows through the shell to eventually reach the inner surface, inducing macroscopic abnormalities such as white spots, conchiolin warts, or abnormal shell growth. Although still observed in France, this disease no longer seems to have a significant impact. In contrast, pathogens such as Marteilia refringens and Bonamia ostreae (Fig. 1) are recognized as major pathogens of the flat oyster and are notifiable to the EU and World Organisation for Animal Health (WOAH). While the first research studies aimed to characterize these parasites and describe their epidemiology (Tab. 1), the acquisition of molecular data later helped to address the phylogenetic relationships between these species and closely related ones and design specific diagnosis tools. Finally, a better understanding of host–parasite interactions and parasite life cycles has enabled disease control and management recommendations to be made for farming activities and restoration or conservation programmes.
 |
Fig. 1 Bonamia ostreae (arrows) in hemocytes of Ostrea edulis (A: histology; B: gill imprint). Marteilia refringens (arrows) in the epithelium of the digestive diverticula in Ostrea edulis (C: histology; D: digestive gland imprint). Pictures by Bruno Chollet / Ifremer. |
3.1 First descriptions of flat oyster parasites in France
In 1968, mortality of flat oysters was reported in Aber Wrach, Brittany, France. The parasite Marteilia refringens was identified as the cause of the disease, known in French as the ‘maladie des abers’ or as marteiliosis. The parasite was first observed in the epithelium of the digestive tract of oysters sampled in Brittany and subsequently in most French flat oyster producing areas (Comps, 1970; Herrbach, 1971). Occasionally, the parasite was also observed in the gills (Grizel et al., 1974). The development of this parasite within the flat oyster is atypical and has been well described: it develops by endogenous divisions along the digestive tract (Grizel et al., 1974; Marteil, 1976). Young plasmodia are mainly found in the epithelium of the oesophagus and stomach (Grizel et al., 1974). Sporulation takes place in the digestive gland tubules and ducts. Propagules are released into the lumen of the digestive tract and shed into the environment in faeces (Audemard et al., 2002; Berthe et al., 2004). Transmission electron microscopy allowed more precise descriptions of the different parasite stages and organelles, including lipidic bodies, refringent bodies in mature stages and haplosporosomes (Longshaw et al., 2001).
Identified as the cause of the maladie hémocytaire de l'huître plate, parasitose hémocytaire or bonamiosis, the parasite Bonamia ostreae was first detected in Ile Tudy, Brittany, France, in summer 1979 (Pichot et al., 1980). It is believed that this parasite was introduced into France and Spain with spat of O. edulis, produced in a hatchery in California, USA (Elston et al., 1986). In histology, 2–3 μm unicellular cells were observed in the cytoplasm of hemocytes and occasionally extracellularly (Pichot et al., 1980). In addition to common cellular organelles such as mitochondria, Golgi apparatus and lipid droplets, ultrastructural examination revealed the presence of dense particles, subsequently named haplosporosomes (Pichot et al., 1980). The movements of farmed stocks contributed to spreading Bonamia ostreae around Brittany (Tigé et al., 1982) and more widely in France and Europe (Pogoda et al., 2019). This parasite has more recently been detected in Ostrea chilensis in New Zealand (Lane et al., 2016).
3.2 Molecular data, taxonomy and phylogeny
Because of the lack of reference genomes for any closely related species and the difficulty of obtaining high numbers of isolated parasites, which are non-culturable, the genomes of Marteilia refringens and Bonamia ostreae are still poorly known. Primers designed in conserved rDNA regions allowed the first partial 18S sequences to be obtained for these parasites (Berthe et al., 2000; Cochennec et al., 2000), confirming that M. refringens belongs to the order Paramyxida and B. ostreae to the order Haplosporida. Subsequently, ITS1-5.8S-ITS2 was partially or entirely characterized, as was IGS for M. refringens (Le Roux et al., 2001; Hill et al., 2010; Kerr et al., 2018; López-Flores et al., 2004). In addition, a few amino acid sequences were characterized for B. ostreae, including two actin genes (López-Flores et al., 2007) and Heat Shock Protein 90 involved in the internalization of the parasite into hemocytes (Prado-Alvarez et al., 2013). More recently, RNA sequencing was carried out from B. ostreae isolated from infected flat oysters (Chevignon et al., 2022). These transcriptomic data were included in a phylogenomic analysis of a set of 229 genes from representative species of all major rhizarian clades, which confirmed the position of B. ostreae within the class Ascetospora (Sierra et al., 2016).
The definition of Marteilia refringens is still under debate. It was initially considered distinct from Marteilia maurini found in the mussels Mytilus galloprovincialis and M edulis (Comps et al., 1982; Auffret and Poder, 1983), but these two Marteilia species were later synonymized under M. refringens based on high identities between their 18S, ITS-1 and IGS sequences. The identification of a polymorphic restriction site within the ITS-1 sequence allowed the description of two genotypes: O-type and M-type, which are predominantly detected in flat oysters and mussels, respectively (Berthe et al., 2000; Le Roux et al., 2001; López-Flores et al., 2004). However, a recent study suggests that O-type and M-type are two distinct species, which were renamed M. refringens and M. pararefringens (Kerr et al., 2018).
3.3 Diagnosis, microscopy and molecular tools
In infected oysters, the macroscopic signs that might be observed include gill perforations or ulcers in cases of B. ostreae and digestive gland discoloration in cases of M. refringens (Comps et al., 1980; Grizel, 1985; Grizel et al., 1974). However, these macroscopic signs are not pathognomonic and most infected oysters appear normal. The examination of histological slides or imprints was used for about twenty years to detect these parasites. In addition, an immunofluorescent technique based on monoclonal antibodies was developed, which offered a level of sensitivity similar to tissue imprints. However, this antibody technique gave unclear results when tested extensively on oysters from Maine, USA (Carnegie and Cochennec-Laureau, 2004). Although a direct monoclonal antibody sandwich immunoassay for the detection of B. ostreae in haemolymph samples of O. edulis was also developed (Cochennec et al., 1992), this tool is no longer available.
Later, sequence data allowed the design of a range of molecular tools including conventional PCR, PCR-RFLP, real-time PCR assays and in situ hybridization (ISH). These tools were shown to be more sensitive than histology or cytology (e.g. Balseiro et al., 2007; Marty et al., 2006; Thébault et al., 2005; López-Florez et al., 2004). Although the first PCR and ISH assays developed were not particularly species specific and could not distinguish closely related parasites, most recent tools show a good specificity (e.g. Robert et al., 2009, Carrasco et al., 2017; Ramilo et al., 2013; EURL, 2022). A duplex Taqman assay was also developed and validated for one-step detection and quantification of the presence of Bonamia spp and/or M. refringens (Canier et al., 2020). This Taqman assay was optimized in order to detect parasite DNA (and RNA for B. ostreae) in environmental samples. These eDNA-based approaches are very promising as they avoid killing oysters, which is important in the context of zone status evaluation, surveillance and restoration programs (Mérou et al., 2020; Mérou, 2021; Mérou et al., 2022).
3.4 Host–parasite interactions and life cycles of parasites
Bonamia ostreae infects all oyster life stages from larvae to adults, although older oysters seem more susceptible to this parasite (Lallias et al., 2008; Arzul et al., 2011; Arzul and Carnegie, 2015). Its transmission is direct from infected to non-infected oysters. The parasite can survive at least two days in seawater depending on the temperature and salinity (Arzul et al., 2009; Mérou et al., 2020). Transmission can occur throughout the year but prevalence and infection levels show a seasonal pattern and peak in late winter–spring (Grizel, 1985; Culloty and Mulcahy, 1996; Arzul et al., 2006; Engelsma et al., 2010) Although methods for in vitro culture of the parasite are lacking, a protocol for parasite purification from infected oysters (Mialhe et al., 1988) has enabled in vitro and in vivo experimental infections (Mialhe et al., 1988; Hervio et al., 1995; Lallias et al., 2008). These infection studies, as well as the availability of flat oysters resistant to bonamiosis, allowed host–parasite interactions to be investigated at cellular and molecular scales (Morga, 2010; Harrang, 2012; Gervais, 2016). This work notably revealed that the parasite activates its own internalization and is able to modulate defence mechanisms, such as the production of reactive oxygen species (ROS) or apoptosis. More recently, a decrease of parasite internalization into hemocytes after an exposure to flat oyster mucus demonstrated the role of the palleal mucus in these host–parasite interactions in vitro (Fernández-Boo et al., 2020).
In contrast to B. ostreae, M. refringens cannot be directly transmitted from infected to non-infected oysters. The involvement of an intermediate host was suspected from early on and the copepod Paracartia grani was identified as a likely candidate (Audemard et al., 2001, 2002; Boyer et al., 2013). Although it is possible to infect copepods with M. refringens from oysters or mussels, trials carried out to infect oysters or mussels from infected copepods have not yet been successful, leaving the parasite cycle unresolved (Audemard, 2001; Carrasco et al., 2008). A congeneric copepod species, Paracartia latisetosa was found infected with M. refringens using PCR and in situ hybridization, suggesting that the parasite may use different intermediate hosts depending on the diversity of zooplankton species present in the ecosystem (Arzul et al., 2014). The transmission of M. refringens generally occurs when the temperature is over 17 °C and prevalence and infection levels show a peak in summer (Audemard, 2001; Carrasco et al., 2007; Boyer, 2012; Arzul et al., 2014). Juveniles and older life stages are known to be susceptible (Grizel, 1985). Recently, an eDNA-based approach allowed parasite DNA to be detected in seawater 20 days after release from the flat oysters (Mérou et al., 2022).
3.5 Disease control and management
Treatments or eradication programs cannot be used to control bivalve diseases in their natural or production environment. Therefore, prevention measures and, particularly, movement restrictions from infected zones are necessary to avoid the spread of bonamiosis and marteiliosis. Stock management measures or relevant husbandry practices allowing producers to decrease the development of these diseases can be adopted to mitigate their impact on flat oyster populations once present in a zone. For example, using suspension culture, decreasing stocking densities and co-cultivation of C. gigas and O. edulis have all been shown to decrease the development of these parasites (Tigé et al., 1984; Grizel, 1985; Le Bec et al., 1991; Lama and Montes, 1993). Farming flat oysters in deeper waters has also been shown to decrease the transmission of M. refringens (Hussenot et al., 2014).
Regarding disinfection, little information is available for these parasites. Contamination of oysters with B. ostreae might be reduced using a peracetic acid bath (0.001% and 0.005%) (Grizel, 1985) or UV above 563.4 mJ/cm2 (Fernández-Boo et al., 2021).
Finally, selective breeding programs have shown that resistance to infection by B. ostreae is heritable (Martin et al., 1992, Culloty et al., 2004). Furthermore, a 54% reduction of parasite prevalence was reported after three generations of selection (Naciri-Graven et al., 1998). The selected oysters then showed a reduced parasite burden and different immune performance compared with non-selected oysters (Morga et al., 2017). These promising results suggest that selecting oysters resistant or tolerant to B. ostreae could contribute to mitigating the impact of the parasite in endemic zones (Holbrook et al., 2021).
3.6 Perspectives for future pathology research
Although many studies have been carried out to improve understanding of marteiliosis and bonamiosis, some key questions remain unresolved. Indeed, the genome of these parasites is still not available. Parasite enrichment protocols have been developed, but the lack of culture method for these parasites prevents us from obtaining their DNA in sufficient quantity and quality. Nevertheless, the recent development of eDNA-based approaches for the detection of B. ostreae and M. refringens (Mérou et al., 2020; Jørgensen et al., 2020; Mérou et al., 2022) will certainly allow their distribution in the ecosystem to be clarified and the health status of flat oyster populations to be monitored without killing oysters. Finally, other pathogens than M. refringens and B. ostreae have also been described in Ostrea edulis (see Colsoul et al., 2021 for a review) and would deserve to be monitored so as to better evaluate their impact on flat oyster populations. The use of wide sequencing approaches including metabarcoding and metagenomics opens new perspectives by making it possible to characterize the pathobiome associated with any shift in health status (Bass et al., 2019).
4 Ecology
Ecological studies on O. edulis in France over the last 50 years are extremely patchy, as shown in Table 1. Only the last 10 years (since 2010) have seen the gradual emergence of studies on its ecology for the purpose of species restoration and these are often only presented in the final reports of regional contracts. The following paragraphs revisit such studies on an international level and attempt to define the most likely chain of causes and processes explaining the decline of flat oysters in France and the difficulties of their self-recovery without human intervention. These aspects will help us identify relevant restoration levers applicable at the regional level.
4.1 Status of the last remnant populations in France
While reefs of the Pacific oyster C. gigas are now very abundant on French coasts, 50 years after the introduction of this species, beds and reefs of the European flat oyster O. edulis are critically endangered in France (Duchene et al., 2015). At the national level, O. edulis is reported in Normandy, New Aquitaine and some Mediterranean lagoons, but is known to be most abundant in Brittany (Hussenot et al., 2014; Duchene et al., 2015). In order to draw up a precise assessment of the conservation status of the last populations present along the Brittany coasts, a georeferenced field monitoring survey was conducted from 2019 to 2021 (Pouvreau et al., 2021b). This study revealed the existence of a small number of significant populations nested in different bays and estuaries, in the heart of areas well-known as the locations of ancient beds and reefs (Fig. 2). A five-level scale was specifically developed for this survey to ecologically classify the conservation status of these populations (Fig. 3). This scale has recently been proposed as a reference for sampling and restoration operations within NORA (zu Ermgassen et al., 2021).
At present, with a few rare exceptions, isolated oysters are mainly observed rolling (stage 0) or fixed but at very low densities (stage 1), sometimes in small very isolated colonies (stage 2), and exceptionally in aggregations of significant density (stage 3). Generally, these observations are made in sheltered, remote and unexploited environments, on bottoms of 0–10 m depth, made up of sandy-muddy sediments, gravel, shell debris or with the oysters directly on boulders. While stage 3 is only exceptionally observed, no populations are classified as stage 4, corresponding to the presumed original reef habitat. Yet, historical evidence and recent restoration operations show that, under optimal conditions, this species is capable of creating biogenic reefs. Given the critical status of observed remaining populations of O. edulis, saving and restoring them has become a high priority in terms of biological conservation (zu Ermgassen et al., 2020). Since 2010, several regional projects have been conducted in France to identify different ways to promote the return of the species and its biogenic habitat (Hussenot et al., 2014; Laguerre, 2014; Cochet et al., 2015; Duchene et al., 2015; Pouvreau et al., 2021a). However, in order to find the best way to guide species restoration efforts, a rigorous analysis of all the causes of its decline is necessary. This analysis is presented below.
 |
Fig. 2 Occurrence of wild beds of flat oysters in Brittany, the colours indicate the stages of their conservation status. For more information, see Pouvreau et al. (2021b). |
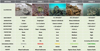 |
Fig. 3 Ecological and conservation stages of beds and reefs of O. edulis observed in Brittany, France. For more information, see Pouvreau et al. (2021b). |
4.2 Revisiting the causes and processes leading to the destruction of flat oyster reefs in France
Cause 1: physical destruction of the habitat and progressive loss of shellfish substrates
The consumption of flat oysters along the French coast is known from the Mesolithic period (Mougne and Dupont, 2013). It was already very important during antiquity and continued during the Middle Ages (Dupont and Rouzeau, 2015), as demonstrated by the large accumulations of shells, known as shell middens found from that period. In addition to their archaeological interest, these mounds bear witness, from an ecological point of view, to the importance of coastal flat oyster populations at this time, which were simply accessible from the foreshore. However, more destructive exploitation of the oyster beds increased steadily from the 16th century onwards with the development of mechanisation (rake fishing, hand dredging, then sailing and motorised fishing) until the habitat was completely eradicated, and the species status very poor. As a testimony of this overexploitation, Table 2 presents some historical quotations reflecting the huge physical degradation of the habitat caused by exploitation in these times. As a result, at the end of the 19th century, a great part of the shells, essential for recruitment of the species, had been completely eradicated from the sea bottom or buried and the physical layer allowing the oysters to settle and grow above the sediment had completely disappeared. This destruction probably indicates a real tipping point in the benthic habitats of the French coast over the last few centuries.
Citations related to the flat oyster fishery explicitly mentioning the physical degradation of the habitat.
Cause 2: land runoff, sedimentation, resuspension and burying
The regular passage of dredging tools on the seabed over centuries was also detrimental on two other levels: it buried remnant live adult oysters and shell material, but also resuspended fine particles which, by being redeposited on the few remaining substrates, then hindered the recruitment of larvae. Suspended matter, carried into the sea by the leaching of land of the catchment areas, which have been increasingly cultivated since the middle of the 20th century in France (Foucher et al., 2021), also contributed to the decline. Although flat oysters are able to inhabit several kinds of environment, as long as there are substrates to attach themselves to, they are less tolerant of muddy sediments than the Pacific oyster. All observations made during monitoring surveys have shown that, unlike the Pacific oyster, O. edulis is systematically absent from over-silted environments or remains only if there are enough clean and elevated substrates to attach to (Pouvreau et al. 2021a). In addition to being used for fixation, these substrates also allow for elevation to within a few centimetres of the sea bottom. Such vertical elevation seems vital to the species, especially in muddy environments, as has been shown in other reef-forming bivalve species (Lenihan, 1999; Lipcius et al., 2015; Colden et al., 2017), particularly because of boundary layer effects (Prandtl, 1926).
Cause 3: role of hydrodynamic dispersion and connectivity
In addition to the quantity and quality of the substrates available on the bottom, the physical circulation of water masses constitutes another determining factor in oyster settlement by imposing strict dispersion patterns on the larvae according to their larval life span and pelagic behaviour. Thus, larval distribution can be very heterogeneous in relation to the hydrodynamics in any given bay, with zones of dilution or zones of concentration. Depending on the location of adult oysters, some areas will therefore be more likely to emit larvae, other areas more likely to receive them, and some may play an intermediate role (Bernard et al., 2014). This non-homogeneous physical forcing plays an essential role in whether a population recovers, depending on its location: a degraded emitting oyster bed has little chance of recovering without external intervention. Identifying dispersal patterns within a marine ecosystem is not an easy task, but there are various tools to address the connectivity issues (Pineda et al., 2009). An indicator of localized dispersion has been specifically developed for restoration purposes in the bay of Brest using a hydrodynamic model called MARS 3D (Petton et al., 2020) and was also validated on Quiberon bay (Stanisière et al., 2016; Pouvreau et al., 2021a). Thanks to this model, certain areas with lower dispersion can be identified. For instance, in the bay of Brest, the model identified low dispersive areas to the south (bay of Daoulas, coves of Poulmic, Roscanvel, Fret and Auberlac'h) and to the north (Keraliou bank) and it is striking to note that all these areas are the same ones known to host the last infralittoral flat oyster populations of the bay (and also the highest supralittoral densities of Pacific oysters). This is therefore very useful for selecting relevant sites for restoration.
Cause 4: reproductive deficit and recruitment variation
The reproductive ecology of the European flat oyster has been the subject of many studies, the first of which date from the beginning of the 20th century when the species supported a flourishing oyster industry (Korringa, 1941: 1946: 1947; Marteil, 1955: 1959: 1960). These studies provide a progressively improving documentation of the effect of environmental parameters (temperature, salinity, current, lunar phase, etc.) on the reproduction of the species while raising numerous questions, most of which remain unanswered. Indeed, since the 1970 s, there has been almost no work in France on O. edulis reproductive ecology, except for two isolated studies (Martin, 1994; Robert et al., 1988). Nevertheless, some studies were carried out on control of reproduction and seed production in hatcheries (see Tab. 1: Robert et al., 1990: 1991: 2017; Gonzalez-Araya, 2011, 2012a, 2012b, 2018; Suquet et al., 2010; Davenel et al., 2010) and natural reproduction of O. edulis (larval abundance and recruitment index) has been monitored each year in the two main bays that still shelter flat oyster beds and where seed collection is performed to supply the local flat oyster industry: the bays of Brest and Quiberon (Cochet, 2004–2020). As an illustration, Figure 4 presents a part of these time series over the last 10 years. These key data show that there are still enough adult oysters in both bays to produce significant larval concentrations that should be enough to allow seed collection for aquaculture as well as relatively simple restoration projects based on natural recruitment. Of course, these larval abundances vary from year to year and within the summer season, depending on environmental conditions, particularly in relation to the temperature of the seawater. This database shows, for instance, that (1) small larvae are observed as soon as the seawater reaches 17 °C, but (2) disappear during the summer if the temperature falls below this threshold, and (3) water temperature must remain above 18 °C for several days to allow full development of larvae and efficient recruitment. Recent warm summers with temperatures close to 20 °C (e.g. 2018, 2019) have been very favourable for the release and development of larvae and their recruitment.
 |
Fig. 4 Temporal variations in abundance of O. edulis larvae in the bays of Brest (Roz station) and Quiberon (Men Er Roué station). The relation with water temperature is indicated by the red strip. Adapted from Pouvreau et al. (2021a). |
Cause 5: effects of biotic interactions on the dynamics of flat oyster populations
The flat oyster is a biogenic species that creates reefs whose three-dimensional complexity influences the biotic interactions of its populations. Among these interactions, predator-prey relationships deserve particular attention, as the more complex the reef structure is, the more it is able to provide spaces providing protection and refuge for young flat oyster recruits (Grabowski, 2004; Barrios-O'Neill et al., 2015). Moreover, the fragmentation of populations on the bottom also increases the attack fronts geometrically and statistically, thus making the oysters more accessible, particularly to crawling predators. Consequently, the progressive degradation of the oyster reef habitat, resulting in the passage from a dense, cohesive reef with a complex three-dimensional structure to sparse individuals with no vertical structure, has probably had the collateral effect of increasing predator pressure (Marteil, 1959) and favouring a vicious circle that leads to an acceleration of the decline of the population to a tipping point beyond which natural recovery is almost impossible. This is why the current level of predation pressure and the way it impedes recovery of the rare wild populations should be examined, even where there is no exploitation. To address this question, a specific experiment was conducted by Pouvreau et al. (2021a) to assess the current level of predation on flat oyster spat in residual populations of flat oysters in the bays of Brest and Quiberon. This study showed that the predation factor is still a major constraint on wild remnant populations: in both sites, the total mortality from predation by oyster drills and sea bream combined reached more than 90% in less than 3 months. Other key biotic interactions affecting the population dynamics of European flat oyster include those related to the parasites M. refringens and B. ostreae, presented in section 3. In the near future, it would also be interesting to study the potential positive effects of epibiosis in the protection of oyster reefs. This has never, to our knowledge, been investigated.
5 Recommendations and perspectives for flat oyster restoration in France
By identifying threats and pressures that prevent recovery of populations as presented in Section 4, but also by using knowledge on genetics and pathology (Sects. 2 and 3), several recommendations for flat oyster restoration in France can clearly be proposed, paying special attention to local and regional constraints. To date, three main active restoration methods have been initiated at a pilot scale in France. These are presented below.
5.1 Active restoration by spat seeding: reintroduction
Seeding with wild spat has been tested in Quiberon bay by the Union of Oyster Farmers of the Bay of Quiberon (‘SOBAIE’) since 2015 (Cochet et al., 2015). Each year, oyster farmers provide 1% of their annual collected spat to be seeded in a former oyster bed located in the centre of Quiberon bay. Before seeding operations, this area is cleaned of crawling predators, especially oyster drills and starfish. Moreover, during all the seeding operations, acoustic repulsive and protective nets are also set up to reduce the predation by sea bream. Regular inspection by scientific dredging and direct diving observation provide information on the growth and density of the seeded juveniles. After 5 years of implementation, this method seems to have been successful, but there remains a strong need for oyster farmers to protect the restored bed from predation.
5.2 Active restoration by shell enrichment: cultch planting
A second approach to restoring ancient flat oyster beds, called ‘oyster cultch restoration', consists of depositing large quantities of mollusc shells, generally from the same (Ostreidae, i.e. Pacific or flat oysters) or related families (Pectinidae, Mytilidae), on the seabed (Beck et al., 2011). This technique is relatively inexpensive and merits to be adapted and applied in France. It is better suited to zones that are calm in terms of hydrodynamics, to avoid the shells being washed away. Additionally, areas that are muddy should be avoided to prevent the added shells from becoming covered in silt and getting buried, which would inhibit recruitment. A first trial of this technique was carried out in the bay of Brest (Pouvreau et al., 2016): 20 bags of 100 litres of Pacific oyster shells were deposited in May 2015 over 50 m2 in the heart of an old flat oyster bed area and these were successfully colonized by successive flat oyster recruitments. In 2017, more than 10 young flat oysters (>1 cm) were observed on each Pacific oyster shell. A similar operation was repeated in July 2021 in the same site and will be evaluated in the coming years. Depositing a bed of limestone before adding shells may improve their efficiency in hydrodynamically turbulent sites.
5.3 Active restoration by laying alternative substrates specifically adapted to the species
To overcome the problem of substrates being buried or washed away, a third approach being tested is to develop eco-designed 3D supports specifically adapted to attract flat oyster larvae and promote settlement. For instance, a limestone concrete reef based on shell fragments has been developed by using Pacific oyster shells (crushed and powdered) mixed with cement according to a specific formulation and set with a specific surface rugosity (Potet et al., 2021). These first artificial reefs dedicated to flat oysters were deployed in the bays of Brest and Quiberon in 2019 and inspected frequently by regular scuba diving to evaluate the natural recruitment, development of young spat, impact of predation, colonization of the reef by other species and long-term resilience of the reef and its population. The long-term evaluation is still in progress, but the first observed densities of flat oysters, level of aggregation and biodiversity were highly encouraging (Pouvreau et al., 2021a). These first results demonstrate the feasibility of active restoration of the species in France using eco-designed 3D supports. New developments are currently in progress concerning this promising method of restoration.
5.4 Genetics and disease management considerations
Whatever the restoration method used, specific attention should be paid to the spat that will be grown on restoration areas. When the origin of the spat is local, it is supposedly adapted to the area and there is no risk of genetic pollution. In this sense, restoration using local spatfall is to be encouraged when still possible. However, when non-local spat is used for restoration, two main questions should be asked before the operations: (1) Is the spat genetically different from the local one? and (2) Is the resource used sufficiently diverse for a sustainable activity?
We know that different genetic clusters coexist in Europe, but also that genetic differences exist at a very local scale. Such genetic differences could be associated with adaptive traits, so moving oysters in a different area may generate maladaptation in the restored site. Moreover, we know that genetic diversity might decline rapidly in farms as well as in the wild, partly due to the specific reproductive biology of the species. For this reason, the level of genetic diversity of a stock to be used for seeding (even a local one) needs to be estimated and optimized before doing the seeding, and should then be followed throughout the restoration program. Genetic markers are available and their use should be included in the restoration programs from their very beginning.
In a similar way, the health status of the restored population should be evaluated by monitoring mortality whenever possible (during diving inspections or scientific sampling) and prevalence of pathogens annually or every two years. Any abnormal mortality should be followed up by oyster sampling and testing using diagnostic tools allowing the detection of known or emerging pathogens. Prevalence of endemic pathogens should be considered as a population health indicator. Measures that contribute to decreasing pathogen transmission and development (e.g. avoiding seeding on the bottom, decreasing density, mixing flat oysters with other species, using spat resistant/tolerant to a pathogen, etc.) could be adopted to mitigate the impact of the pathogen on the oyster population.
In addition, in the context of an active restoration by spat seeding and reintroduction, the spat should also be tested for the presence of pathogens prior to seeding. Lastly, in the culch planting scenario, the mollusc shells should be carefully cleaned in order to avoid the introduction of associated species including pathogenic agents present on the surface of the shells or in the remaining tissues.
6 Concluding remarks
A large number of research projects and articles have been devoted to the native European flat oyster Ostrea edulis. This is notably the case in France, which remains the main producing country in Europe. Most of these projects have targeted aquaculture applications while and ecology of the species in its natural habitat has been neglected until recent years. Fortunately, despite their massive overall decline, wild beds can still be found in France, notably in Brittany. This provides a unique opportunity to implement more integrated research projects uniting the different disciplines covered in this review article, so that restoration programs can benefit coastal ecosystems, marine biodiversity and sustainable aquaculture. Recent results of restoration programs in Brittany are very encouraging, so we hope that this review article will contribute to informing and inspiring French stakeholders so that local flat oyster beds will recover in the coming decades, under the framework of the Native Oyster Restoration Alliance (NORA) and as part of the UN Decade for Ecosystem Restoration.
Acknowledgements
Some of the results presented in this article were funded by the European Maritime and Fisheries Fund (FEAMP project no. 17/2215675) within the FOREVER project.
References
- Alleway H, Connell SD. 2015. Loss of an ecological baseline through the eradication of oyster reefs from coastal ecosystems and human memory. Conserv Biol 29: 795–804. [CrossRef] [PubMed] [Google Scholar]
- Anonyme. 2021. Plan Aquacultures d'Avenir, 2021–2027. Rapport Ministère de la mer. 83 p. [Google Scholar]
- Arzul I, Carnegie RB. 2015. New Perspective on The Haplosporidian Parasites of Molluscs. J Invertebr Pathol 131: 32–42. [CrossRef] [PubMed] [Google Scholar]
- Arzul I, Chollet B, Boyer S, Bonnet D, Gaillard J, Baldi Y, Robert M, Joly JP, Garcia C, Bouchoucha M. 2014. Contribution to the understanding of the cycle of the protozoan parasite Marteilia refringens. Parasitology 141: 227–240. [CrossRef] [PubMed] [Google Scholar]
- Arzul I, Gagnaire B, Bond C, Chollet B, Morga B, Ferrand S, Robert M, Renault T. 2009. Effects of temperature and salinity on the survival of Bonamia ostreae, a parasite infecting flat oysters Ostrea edulis. Dis Aquat Org 85: 67–75. [CrossRef] [PubMed] [Google Scholar]
- Arzul I, Langlade A, Chollet B, Robert M, Ferrand S, Omnes E, Lerond S, Couraleau Y, Joly JP, François C, Garcia C. 2011. Can the protozoan parasite Bonamia ostreae infect larvae of flat oysters Ostrea edulis? Vet Parasitol 179: 69–76. [CrossRef] [PubMed] [Google Scholar]
- Arzul I, Miossec L, Blanchet E, Garcia C, Francois C, Joly JP. 2006. Bonamia ostreae and Ostrea edulis : a stable host-parasite system in France? In: Proceedings of the 11th Symposium of the International Society for Veterinary Epidemiology and Economics (ISVEE), Cairns, Queensland, Australia, 6-11 August 2006. https://archimer.ifremer.fr/doc/00000/6381/ [Google Scholar]
- Audemard C, Barnaud A, Collins CM, Le Roux F, Sauriau PG, Coustau C, Blachier P, Berthe FCJ. 2001. Claire ponds as an experimental model for Marteilia refringens life-cycle studies: new perspectives. J Exp Mar Biol Ecol 257: 87–108. [CrossRef] [PubMed] [Google Scholar]
- Audemard C, Le Roux F, Barnaud A, Collins C, Sautour B, Sauriau PG, De Montaudouin X, Coustau C, Combes C, Berthe F. 2002. Needle in a haystack: involvement of the copepod Paracartia grani in the life-cycle of the oyster pathogen Marteilia refringens. Parasitology 124: 315–323. [CrossRef] [PubMed] [Google Scholar]
- Audemard C. 2001. Stratégie d'utilisation de différentes espèces animales par le parasite Marteilia refringens pour assurer son cycle biologique. PhD Thesis, Université de Perpignan, 186 p. https://archimer.ifremer.fr/doc/00107/21850/ [Google Scholar]
- Auffret M, Poder M. 1983. Recherches sur Marteilia maurini, parasite de Mytilus edulis sur les côtes de Bretagne nord. (Studies on Marteilia maurini, parasite of Mytilus edulis from the north coasts of Brittany). Rev Trav Inst Pêche Marit 47: 105–109 (In French with English abstract). [Google Scholar]
- Balseiro P, Montes A, Ceschia G, Gestal C, Novoa B, Figueras A. 2007. Molecular epizootiology of the European Marteilia spp., infecting mussels (Mytilus galloprovincialis and M edulis) and oysters (Ostrea edulis): an update. Bull Eur Assoc Fish Pathol 27: 148–156. [Google Scholar]
- Barrios-O'Neill D, Dick JTA, Emmerson MC, Ricciardi A, Macisaac HJ. 2015. Predator-free space, functional responses and biological invasions. Funct Ecol 29: 377–384. [CrossRef] [Google Scholar]
- Bass D, Stentiford GD, Wang HC, Koskella B, Tyler CR. 2019. The pathobiome in animal and plant diseases. Trends Ecol Evol 34: 996–1008. [CrossRef] [PubMed] [Google Scholar]
- Baud JP, Gérard A, Naciri-Graven Y. 1997. Comparative growth and mortality of Bonamia ostreae-resistant and wild flat oysters, Ostrea edulis, in an intensive system. I. First year of experiment. Mar Biol 130: 71–79. [CrossRef] [Google Scholar]
- Beaumont A, Garcia MT, Hönig S, Low P. 2006. Genetics of Scottish populations of the native oyster, Ostrea edulis: gene flow, human intervention and conservation. Aquat Living Resour 19: 389–402. [CrossRef] [EDP Sciences] [Google Scholar]
- Bean T, Tanguy A, Penaloza C, Gundappa M, Boutet I, Houston RD, Macqueen DL, Boudry P. 2022. Two parallel chromosome-level reference genomes to support restoration and aquaculture of European flat oyster Ostrea edulis. Evolut Appl 15: 1709–1712. [CrossRef] [Google Scholar]
- Beck MW, Brumbaugh RD, Airoldi L, Carranza A, Coen LD, Crawford C, Defeo O, Edgar GJ, Hancock B, Kay MC, Lenihan HS, Luckenbach MW, Toropova CL, Zhang G, Guo X. 2011. Oyster reefs at risk and recommendations for conservation, restoration, and management. BioScience 61: 107–116. [CrossRef] [Google Scholar]
- Bédier E, Baud J-P, Cochennec N, Cornette F, Gérard A, Goyard E, Joly J-P, Kopp J, Langlade A, Le Coguic Y, Mazurié J, Nourry M. 2001. L'amélioration des performances de l'huître plate européenne Ostrea edulis par la sélection génétique. Actes Journées Conchylicoles, Ifremer Nantes, 3-4 avril 2001. https://archimer.ifremer.fr/doc/00000/3274 [Google Scholar]
- Bernard I, Duchene J, Hily C, Lazure P, Pouvreau S, Boudry P. 2014. Étude de la connectivité potentielle en rade de Brest : application au cas de l'huître plate, Ostrea edulis. Rapport de contrat, 24p. https://archimer.ifremer.fr/doc/00251/36233/ [Google Scholar]
- Bernatchez S, Xuereb A, Laporte M, Benestan L, Steeves R, Laflamme M, Bernatchez L, Mallet MA. 2019. Seascape genomics of Eastern Oyster (Crassostrea virginica) along the Atlantic Coast of Canada. Evol Appl 12: 587–609. [CrossRef] [PubMed] [Google Scholar]
- Berthe FCJ, Le Roux F, Adlard RD, Figueras A. 2004. Marteiliosis in molluscs: a review. Aquat Living Resour 17: 433–448. [CrossRef] [EDP Sciences] [Google Scholar]
- Berthe FCJ, Le Roux F, Peyretaillade E, Peyret P, Rodriguez D, Gouy M, Vivarès CP. 2000. Phylogenetic analysis of the small subunit ribosomal RNA of Marteilia refringens validates the existence of Phylum Paramyxea (Desportes and Perkins 1990). J Eukaryot Microbiol 47: 288–293. [CrossRef] [PubMed] [Google Scholar]
- Bertolini C, Pastres R. 2022. Identifying knowledge gaps for successful restorative aquaculture of Ostrea edulis: a bibliometric analysis. Open Res Europe 1: 103. [CrossRef] [Google Scholar]
- Bierne N, Gagnaire P-A, David P. 2013. The geography of introgression in a patchy environment and the thorn in the side of ecological speciation. Curr Zool 59: 72–86. [CrossRef] [Google Scholar]
- Blanc F, Pichot P, Attard J. 1986. Genetic variability in the European oyster, Ostrea edulis: geographic variation between local French stocks. Aquaculture 57: 362–363. [CrossRef] [Google Scholar]
- Boudry P, Heurtebise S, Collet B, Cornette F, Gérard A. 1998. Differentiation between populations of the Portuguese oyster, Crassostrea angulata (Lamark) and the Pacific oyster, Crassostrea gigas (Thunberg), revealed by mtDNA RFLP analysis. J Exp Mar Biol Ecol 226: 279–291. [CrossRef] [Google Scholar]
- Boutet I, Alves Monteiro HJ, Baudry L, Takeuchi T, Bonnivard E, Billoud B, Farhat S, Gonzales-Haraya R, Salaun B, Andersen A, Toullec J-Y, Lallier FH, Flot J-F, Guiglielmoni N, Guo X, Li C, Allam B, Pales-Espinosa E, Hemmer-Hansen J, Moreau P, Marbouty M, Koszul R, Tanguy A. 2022. Chromosomal assembly of the flat oyster (Ostrea edulis L.) genome as a new genetic resource for aquaculture. 1730. Evolut Appl 15: 1730–1748. [CrossRef] [Google Scholar]
- Boyer S, Chollet B, Bonnet D, Arzul I. 2013. New evidence for the involvement of Paracartia grani (Copepoda, Calanoida) in the life cycle of Marteilia refringens (Paramyxea). Int J Parasitol 43: 1089–1099. [CrossRef] [PubMed] [Google Scholar]
- Boyer S. 2012. Ecologie du copépode calanoïde Paracartia grani. Implication dans le cycle de vie du parasite Marteilia refringens dans la lagune de Thau. PhD Thesis, Université de Montpellier II, 310 p. https://archimer.ifremer.fr/doc/00118/22931/ [Google Scholar]
- Canier L, Dubreuil C, Noyer M, Serpin D, Garcia C, Arzul I. 2020. A new multiplex real-time PCR assay to improve the diagnosis of shellfish regulated parasites of the genus Marteilia and Bonamia. Prev Vet Med 183: 105126 (8p.) [CrossRef] [PubMed] [Google Scholar]
- Carnegie RB, Cochennec-Laureau N. 2004. Microcell parasites of oysters: recent insights and future trends. Aquat Living Resour 17: 519–528. [CrossRef] [EDP Sciences] [Google Scholar]
- Carpentier C, Mille D, Oudot G, Bodin P, Dhommée R, Phiphak B, Boulette M. 2019. Guide technique de l'élevage d'huîtres plates dans le bassin de Marennes-Oléron. Rapport du Programme IDCEP, 36 p. [Google Scholar]
- Carrasco N, Arzul I, Chollet B, Robert M, Joly JP, Furones MD, Berthe FCJ. 2008. Comparative experimental infection of the copepod Paracartia grani with Marteilia refringens and Marteilia maurini. J Fish Dis 31: 497–504. [CrossRef] [PubMed] [Google Scholar]
- Carrasco N, López-Flores I, Alcaraz M, Furones MD, Berthe FCJ, Arzul I. 2007. Dynamics of the parasite Marteilia refringens (Paramyxea) in Mytilus galloprovincialis and zooplankton populations in Alfacs Bay (Catalonia, Spain). Parasitology 134: 1541–1550. [CrossRef] [PubMed] [Google Scholar]
- Carrasco N, Voorbergen-Laarman M, Lacuesta B, Furones D, Engelsma MY. 2017. Application of a competitive real time PCR for detection of Marteilia refringens genotype “O” and ”M” in two geographical locations: the Ebro Delta, Spain and the Rhine-Meuse Delta, the Netherlands. J Invertebr Pathol 149: 51–55. [CrossRef] [PubMed] [Google Scholar]
- Chevignon G, Serpin D, Dotto-Maurel A, Chollet B, Arzul I. 2022. De novo transcriptome assembly and analysis of the flat oyster pathogenic protozoa Bonamia ostreae. Front Cell Infect Microbiol doi: 10.3389/fcimb.2022.921136 [PubMed] [Google Scholar]
- Cochennec N, Hervio D, Panatier B, Boulo V, Mialhe E, Rogier H, Grizel H, Paolucci F. 1992. A direct monoclonal antibody sandwich immunoassay for detection of Bonamia ostreae (Acetospora) in hemolymph samples of the flat oyster Ostrea edulis (Mollusca: Bivalvia). Dis Aquat Org 12: 129–134. [CrossRef] [Google Scholar]
- Cochennec N, Le Roux F, Berthe F, Gerard A. 2000. Detection of Bonamia ostreae based on small subunit ribosomal probe. J Invertebr Pathol 76: 26–32. [CrossRef] [PubMed] [Google Scholar]
- Cochet H, Pouvreau S, Tanguy L, Kerisit C. 2015. Estimation, Préservation, Reconstitution des bancs sauvages d'huîtres plates en Baie de Quiberon. Rapport de contrat, 24 p. [Google Scholar]
- Cochet H. 2004–2020. Bilans annuels du suivi de la reproduction de l'huître plate en Baie de Quiberon. Annual reports from 2004 to 2020. https://sites.google.com/view/cochet-environnement/accueil [Google Scholar]
- Colden AM, Latour RJ, Lipcius RN. 2017. Reef height drives threshold dynamics of restored oyster reefs. Mar Ecol Prog Ser 582: 1–13. [CrossRef] [Google Scholar]
- Colsoul B, Boudry P, Pérez‐parallé ML, Bratoš Cetinić A, Hugh‐Jones T, Arzul I, Mérou N, Wegner KM, Peter C, Merk V, Pogoda B. 2021. Sustainable large‐scale production of European flat oyster (Ostrea edulis) seed for ecological restoration and aquaculture: a review. Rev Aquac 13: 1423–1468. [CrossRef] [Google Scholar]
- Comps M, Pichot Y, Papagianni P. 1982. Recherche sur Marteilia maurini n. sp. parasite de la moule Mytilus galloprovincialis Lmk. Rev Trav Inst Pêche Marit 45: 211–214. [Google Scholar]
- Comps M, Tigé G, Grizel H. 1980. Étude ultrastructurale d'un protiste parasite de l'huître Ostrea edulis. C R Acad Sci Paris Série D 290: 383–384. [Google Scholar]
- Comps M. 1970. Observations sur les causes d'une mortalité anormale des huîtres plates dans le bassin de Marennes. Rev Trav Inst Pêche Marit 34: 317–326. [Google Scholar]
- Culloty SC, Cronin MA, Mulcahy MF. 2001. An investigation into the relative resistance of Irish flat oysters Ostrea edulis L. to the parasite Bonamia ostreae (Pichot et al., 1980). Aquaculture 199: 229–244. [Google Scholar]
- Culloty SC, Cronin MA, Mulcahy MF. 2004. Potential resistance of a number of populations of the oyster Ostrea edulis to the parasite Bonamia ostreae. Aquaculture 237: 41–58. [Google Scholar]
- Culloty SC, Mulcahy MF. 1996. Season-, age-, and sex-related variations in the prevalence of bonamiasis in flat oyster (Ostrea edulis L.) on the south coast of Ireland. Aquaculture 144: 53–63 [CrossRef] [Google Scholar]
- Da Costa F, Gonzales-Araya R, Robert R. Using combinations of microalgae to condition European flat oyster (Ostrea edulis) broodstock and feed the larvae: effects on reproduction, larval production and development. Aquaculture 568: 739302. [Google Scholar]
- Danic-Tchaleu G, Heurtebise S, Morga B, Lapègue S. 2011. Complete mitochondrial DNA sequence of the European flat oyster Ostrea edulis confirms Ostreidae classification. BMC Res Notes 4: 400. [CrossRef] [PubMed] [Google Scholar]
- Davenel A, Gonzalez R, Suquet M, Quellec S, Robert R. 2010. Individual monitoring of gonad development in the European flat oyster Ostrea edulis by in vivo magnetic resonance imaging. Aquaculture 307: 165–169. [CrossRef] [Google Scholar]
- DeWoody JA, Harder AM, Mathur S, Willoughby J.R. 2021. The long‐standing significance of genetic diversity in conservation. Mol Ecol 30: 4147–4154. [CrossRef] [PubMed] [Google Scholar]
- Diaz-Almela E, Boudry P, Launey S, Bonhomme F, Lapègue S. 2004. Reduced female gene flow in the European flat oyster Ostrea edulis. J Hered 95: 510–516. [CrossRef] [PubMed] [Google Scholar]
- Duchene J, Bernard I, Pouvreau S. 2015. Vers un retour de l'huître indigène en rade de Brest. Espèces 16: 51–57. [Google Scholar]
- Dupont C, Rouzeau N. 2015. Des exploitations intensives d'huîtres pendant l'Antiquité et le Moyen âge sur le littoral atlantique français : L'exemple de Beauvoir-sur-Mer (Vendée). Anthropozoologica 50: 109–122. [CrossRef] [Google Scholar]
- Elston RA, Farley CA, Kent ML. 1986. Occurrence of bonamiosis in European flat oysters Ostrea edulis in North America. Dis Aquat Org 2: 49–54. [CrossRef] [Google Scholar]
- Engelsma MY, Kerkhoff S, Roozenburg I, Haenen OLM, van Gool A, Sistermans W, Wijnhoven S, Hummel H. 2010. Epidemiology of Bonamia ostreae infecting European flat oyster Ostrea edulis from Lake Grevelingen, The Netherlands. Mar Ecol Prog Ser 409: 131–142. [CrossRef] [Google Scholar]
- EURL 2022. https://www.eurl-mollusc.eu/content/download/137231/file/B.ostreae%26B.exitiosa%20_TaqmanRealTimePCR_editionN%C2%B01.pdf [Google Scholar]
- Fallet M, Luquet E, David P, Cosseau C. 2020. Epigenetic Inheritance and Intergenerational Effects in Mollusks. Gene 729: 144166. [CrossRef] [PubMed] [Google Scholar]
- Fernández-Boo S, Gervais O, Prado-Alvarez M, Chollet B, Claverol S, Lecadet C, Dubreuil C, Arzul I . 2020. Is pallial mucus involved in Ostrea edulis defenses against the parasite Bonamia ostreae? J Invertebr Pathol 169: 107259. [Google Scholar]
- Fernández-Boo S, Provot C, Lecadet C, Stavrakakis C, Papin M, Chollet B, Auvray JF, Arzul I. 2021. Inactivation of marine bivalve parasites using UV-C irradiation: examples of Perkinsus olseni and Bonamia ostreae. Aquac Rep 21: 100859. [Google Scholar]
- Foucher A, Evrard O, Cerdan O, Chabert C, Lefèvre I, Vandromme R, Salvador-Blanes S. 2021. Deciphering human and climatic controls on soil erosion in intensively cultivated landscapes after 1950 (Loire Valley, France). Anthropocene 34: 1–11. [Google Scholar]
- Gervais O. 2016. Etude cellulaire et moléculaire de l'apoptose chez l'huître plate Ostrea edulis en réponse au parasite Bonamia ostreae / Cellular and molecular study of apoptosis in the flat oyster Ostrea edulis in response to Bonamia ostreae. PhD Thesis, Université de La Rochelle, 240 p. [Google Scholar]
- Glize P, Meneur C. 2018. Diversification conhylicole en baie de Bourgneuf : Potentialités de l'élevage au large (offshore). Rapport de Contrat DICOBAIE, SMIDAP, 22p. [Google Scholar]
- Gonzalez-Araya R, Lebrun L, Quere C, Robert R. 2012b. The selection of an ideal diet for Ostrea edulis (L.) broodstock conditioning (part B). Aquaculture 362–363: 55–66. [Google Scholar]
- Gonzalez-Araya R, Mingant C, Petton B, Robert R. 2012a. Influence of diet assemblage on Ostrea edulis broodstock conditioning and subsequent larval development. Aquaculture 364-365: 272–280. [Google Scholar]
- Gonzalez-Araya R, Queau I, Quere C, Moal J, Robert R. 2011. A physiological and biochemical approach to selecting the ideal diet for Ostrea edulis (L.) broodstock conditioning (part A). Aquac Res 42: 710–726. [CrossRef] [Google Scholar]
- Gonzalez-Araya R, Robert R. 2018. Larval development and fatty acid composition of Ostrea edulis (L.) fed four different single diets from conditioning to pre-settlement. Aquac Res 49: 1768–1781. [CrossRef] [Google Scholar]
- Grabowski JH. 2004. Habitat complexity disrupts predator-prey interactions but not the trophic cascade on oyster reefs. Ecology 85: 995–1004. [CrossRef] [Google Scholar]
- Grizel H, Comps M, Bonami JR, Cousserans F, Duthoit JL, LePennec MA. 1974. Recherche sur l'agent de la maladie de la glande digestive de Ostrea edulis Linné. Bull Inst Pêches Mar 240: 7–30. [Google Scholar]
- Grizel H, Héral M. Introduction into France of the Japanese oyster (Crassostrea gigas). J Conseil 47: 399–403. [Google Scholar]
- Grizel H. 1985. Etudes des récentes épizooties de l'huître plate Ostrea edulis L. et de leur impact sur l'ostréiculture bretonne. Thèse de doctorat, Université des Sciences et Techniques de Languedoc, Montpellier, France, 232 p. https://archimer.ifremer.fr/doc/00000/2581/ [Google Scholar]
- Guillet J, Guillet R. 2008. L'ostréiculture en Bretagne : De 1850 à nos jours. Coop Breizh Press. 320 p. [Google Scholar]
- Gundappa MK, Peñaloza C, Regan T, Boutet I, Tanguy A, Houston RD, Bean TP, Macqueen DJ. 2022. Chromosome level reference genome for European flat oyster (Ostrea edulis L.). Evol Appl 15: 1713–1729. [CrossRef] [PubMed] [Google Scholar]
- Gutierrez AP, Turner F, Gharbi K, Talbot R, Lowe NR, Peñaloza C, McCullough M, Prodöhl PA, Bean TP, Houston RD. 2017. Development of a Medium Density Combined-Species SNP Array for Pacific and European Oysters (Crassostrea gigas and Ostrea edulis). G3 Genes Genom Genet 7: 2209–2218. [Google Scholar]
- Harrang E, Heurtebise S, Faury N, Robert M, Arzul I, Lapègue S. 2015. Can survival of European flat oysters following experimental infection with Bonamia Ostreae be predicted using QTLs? Aquaculture 448: 521–530. [CrossRef] [Google Scholar]
- Harrang E, Lapègue S, Morga B, Bierne N. 2013. A high load of non-neutral amino-acid polymorphisms explains high protein diversity despite moderate effective population size in a marine bivalve with sweepstakes reproduction. G3 Genes Genom Genet 3: 333–341. [Google Scholar]
- Harrang E. 2012. Apport des informations moléculaires et cellulaires pour la caractérisation de la résistance de l'huître plate européenne vis-à-vis de la bonamiose, et pour la détection de signatures de la sélection naturelle. PhD thesis, Université de La Rochelle, 322 p. https://archimer.ifremer.fr/doc/00127/23785/ [Google Scholar]
- Haure J, Penisson C, Bougrier S, Baud JP. 1998. Influence of temperature on clearance and oxygen consumption rates of the flat oyster Ostrea edulis: determination of allometric coefficients. Aquaculture 169: 211–224. [CrossRef] [Google Scholar]
- Haure J. 1999. Testage de souches d'huîtres plates Ostrea edulis tolérantes à Bonamia ostreae: élevage contrôlé, écophysiologie et modélisation de la croissance. PhD thesis, Ecole Pratique des Hautes Etudes, 118 p. https://w3.ifremer.fr/archimer/doc/00108/21961/19556.pdf [Google Scholar]
- Hawkins AJS, Day A, Gerard A, Naciri-Graven Y, Ledu C, Bayne B, Héral M. 1994. A genetic and metabolic basis for faster growth among triploids induced by blocking meiosis I but not meiosis II in the larviparous European flat oyster, Ostrea edulis L. J Exp Mar Biol Ecol 184: 21–40. [CrossRef] [Google Scholar]
- Hedgecock D, Launey S, Pudovkin AI, Naciri Y, Lapègue S, Bonhomme F. 2007. Small effective number of parents (Nb) inferred for a naturally spawned cohort of juvenile European flat oysters Ostrea edulis. Mar Biol 150: 1173–1182. [CrossRef] [Google Scholar]
- Hedgecock D, Pudovkin A.I. 2011. Sweepstakes reproductive success in highly fecund marine fish and shellfish: a review and commentary. Bull Mar Sci 87: 971–1002. [CrossRef] [Google Scholar]
- Herrbach B. 1971. Sur une affection parasitaire de la glande digestive de l'huître plate, Ostrea edulis Linne. Rev Trav Inst. Pêche Marit 35: 79–87. [Google Scholar]
- Hervio D, Bachère E, Boulo V, Cochennec N, Vuillemin V, Le Coguic Y, Cailletaux G, Mazurie J‚ Mialhe E. 1995. Establishment of an experimental infection protocol for the flat oyster, Ostrea edulis, with the intrahaemocytic protozoan parasite, Bonamia ostreae: application in the selection of parasite-resistant oysters. Aquaculture 132: 183–194. [CrossRef] [Google Scholar]
- Hill KM, Carnegie RB, Aloui-Bejaoui N, Gharsalli RE, White DM, Stokes NA, Burreson EM. 2010. Observation of a Bonamia sp. infecting the oyster Ostrea stentina in Tunisia, and a consideration of its phylogenetic affinities. J Invertebr Pathol 103: 179–185. [CrossRef] [PubMed] [Google Scholar]
- Holbrook Z, Bean TP, Lynch SA, Hauton C. 2021. What do the terms resistance, tolerance, and resilience mean in the case of Ostrea edulis infected by the haplosporidian parasite Bonamia ostreae. J Invertebr Pathol 182: 107579. [CrossRef] [PubMed] [Google Scholar]
- Hussenot M, Pouvreau S, Duchene J, Freulon H, Arzul I, Lapègue S. 2014. Synthèse PERLE. Programme d'Expérimentation et de recherche sur L'huître plate Ostrea edulis. 25 p. https://archimer.ifremer.fr/doc/00249/36060/ [Google Scholar]
- Jaziri H. 1990. Variations génétiques et structuration biogéographique chez un bivalve marin : l'huître plate Ostrea edulis L.: PhD Thesis, Montpellier, 148 p. [Google Scholar]
- Johannesson K, Rödström EM, Aase H. 1989. Low genetic variability in Scandinavian populations of Ostrea edulis L. − Possible causes and implications. J Exp Mar Biol Ecol 128: 177–190. [CrossRef] [Google Scholar]
- Jørgensen von Gersdorff F L, S Nielsen JW, Villadsen MK, Vismann B, Dalvin S, Mathiessen H, Madsen L, Kania PW, Buchmann K. 2020. A non-lethal method for detection of Bonamia ostreae in flat oyster (Ostrea edulis) using environmental. DNA Sci Rep 10: 16143. [Google Scholar]
- Joubin L. 1908. La carte des mollusques comestibles des côtes de France. Ann Géogr 93: 197–204. [CrossRef] [Google Scholar]
- Kerr R, Ward GM, Stentiford GD, Alfjorden A, Mortensen S, Bignell JP, Feist SW, Villalba A, Carballal MJ, Cao A, Arzul I, Ryder D, Bass D. 2018. Marteilia refringens and Marteilia pararefringens sp. nov. are distinct parasites of bivalves and have different European distributions. Parasitology 145: 1483–1492. [CrossRef] [PubMed] [Google Scholar]
- Korringa P. 1941. Experiments and observations on swarming, pelagic life and setting in the European flat oyster, Ostrea edulis. Arch Néerlandaises Sci Exactes Nat 5: 1–241. [Google Scholar]
- Korringa P. 1946. A revival of natural oyster beds? Nature 4017: 586–587. [CrossRef] [Google Scholar]
- Korringa P. 1947. Relations between the moon and periodicity in the breeding of marine animals. Ecol Soc Am 17: 347–381. [Google Scholar]
- Laguerre H. 2014. Gisement d'huîtres plates de l'Odet : caractérisation écologique du banc de Perennou. Rapport Agro Campus. Compte rendu d'étude, 34 p. [Google Scholar]
- Lallias D, Arzul I, Heurtebise S, Ferrand S, Chollet B, Robert M, Beaumont AR, Boudry P, Morga B, Lapègue S. 2008. Bonamia ostreae-induced mortalities in one-year old European flat oysters Ostrea edulis: experimental infection by cohabitation challenge. Aquat Living Resour 21: 423–439. [CrossRef] [EDP Sciences] [Google Scholar]
- Lallias D, Beaumont AR, Haley CS, Boudry P, Heurtebise S, Lapègue S. 2007. A first-generation genetic linkage map of the European flat oyster Ostrea edulis (L.) based on AFLP and microsatellite markers. Anim Genet 38: 560–568. [CrossRef] [PubMed] [Google Scholar]
- Lallias D, Boudry P, Lapègue S, King JW, Beaumont AR. 2010a. Strategies for the retention of high genetic variability in European flat oyster (Ostrea edulis) restoration programmes. Conserv Genet 11: 1899–1910. [CrossRef] [Google Scholar]
- Lallias D, Gomez-Raya L, Haley CS, Arzul I, Heurtebise S, Beaumont AR, Boudry P, Lapègue S. 2009b. Combining two-stage testing and interval mapping strategies to detect QTL for resistance to bonamiosis in the European flat oyster Ostrea edulis. Mar Biotechnol 11: 570–584. [CrossRef] [PubMed] [Google Scholar]
- Lallias D, Stockdale R, Boudry P, Beaumont AR, Lapègue S, 2009a. Characterization of 27 microsatellite loci in the European flat oyster Ostrea edulis. Mol Ecol Res 9: 960–963. [CrossRef] [Google Scholar]
- Lallias D, Taris N, Boudry P, Bonhomme F, Lapègue S. 2010b. Variance in the reproductive success of flat oyster Ostrea edulis L. assessed by parentage analyses in natural and experimental conditions. Genet Res 92: 175–187. [CrossRef] [PubMed] [Google Scholar]
- Lama A, Montes J. 1993. Influence of depth of culture in the infection of the European flat oyster (Ostrea edulis L.) by Bonamia ostreae. Bull Eur Assoc Fish Pathol 13: 17–20. [Google Scholar]
- Lane HS, Webb SC, Duncan J. 2016. Bonamia ostreae in the New Zealand oyster Ostrea chilensis: a new host and geographic record for this haplosporidian parasite. Dis Aquat Org 118: 55–63. [CrossRef] [PubMed] [Google Scholar]
- Lapègue S, Harrang E, Heurtebise S, Flahauw E, Donnadieu C, Gayral P, Ballenghien M, Genestout L, Barbotte L, Mahla R, Haffray P, Klopp C. 2014. Development of SNP genotyping arrays in two shellfish species. Mol Ecol Res 14: 820–830. [CrossRef] [Google Scholar]
- Lapègue S, Reisser C, Harrang E, Heurtebise S, Bierne B. 2023. Genetic parallelism between European flat oyster populations at the edge of their natural range. Evol Appl 16: 393–407. [CrossRef] [PubMed] [Google Scholar]
- Launey S, Barre M, Gerard A, Naciri-Graven Y. 2001. Population bottleneck and effective size in Bonamia ostreae-resistant populations of Ostrea edulis as inferred by microsatellite markers. Genet Res 78: 259–270. [CrossRef] [Google Scholar]
- Launey S, Ledu C, Boudry P, Bonhomme F, Naciri-Graven Y. 2002. Geographic structure in the European flat oyster (Ostrea edulis L.) as revealed by microsatellite polymorphism. J Hered 93: 331–338. [CrossRef] [PubMed] [Google Scholar]
- Le Bec C, Mazurie J, Cochennec N, Le Coguic Y. 1991. Influence of Crassostrea gigas mixed with Ostrea edulis on the incidence of Bonamia disease. Aquaculture 93: 263–271. [CrossRef] [Google Scholar]
- Le Roux F, Lorenzo G, Peyret P, Audemard C, Figueras A, Vivarès C, Gouy M, Berthe F. 2001. Molecular evidence for the existence of two species of Marteilia in Europe. J Eukaryot Microbiol 48: 449–454. [CrossRef] [PubMed] [Google Scholar]
- Leitao A, Chaves R, Santos S, Guedes Pinto FH, S Boudry P. 2004. Restriction enzyme digestion chromosome banding in Crassostrea and Ostrea species: comparative karyological analysis within Ostreidae. Genome 47: 781–788. [CrossRef] [PubMed] [Google Scholar]
- Lenihan HS. 1999. Physical-biological coupling on oyster reefs: How habitat structure influences individual performance. Ecol Monogr 69: 251–275. [Google Scholar]
- Levasseur O. 2006. Histoire de l'huître en Bretagne. Skol Vreizh Press, 90 p. [Google Scholar]
- Li X, Bai Y, Dong Z, Xu C, Liu S, Yu H, Kong L, Li Q. 2023. Chromosome-level genome assembly of the European flat oyster (Ostrea edulis) provides insights into its evolution and adaptation. Comp Biochem Physiol Part D Genomics Proteomics 45: 101045. [Google Scholar]
- Lipcius RN, Burke RP, Mcculloch DN, Schreiber SJ, Schulte DM, Seitz RD, Shen J, Willis TJ, Drew JA. 2015. Overcoming restoration paradigms: value of the historical record and metapopulation dynamics in native oyster restoration Front Mar Sci 2: 1–15. [CrossRef] [Google Scholar]
- Longshaw M, Feist SW, Matthews RA, Figueras A. 2001. Ultrastructural characterisation of Marteilia species (Paramyxea) from Ostrea edulis, Mytilus edulis and Mytilus galloprovincialis in Europe. Dis Aquat Org 44: 137–142. [CrossRef] [PubMed] [Google Scholar]
- López-Flores I, de la Herrán FR, Garrido-Ramos MA, Navas JI, Ruiz Rejón F M. 2004. The molecular diagnosis of Marteilia refringens and differentiation between Marteilia strains infecting oysters and mussels based on the rDNA IGS sequence. Parasitology 129: 411–419. [CrossRef] [PubMed] [Google Scholar]
- López-Flores L, Suarez-Santiago VN, Longet D, Saulnier D, Chollet B, Arzul I. 2007. Characterization of actin genes in Bonamia ostreae and their application to phylogeny of the Haplosporidia. Parasitology 134: 1941–1948. [CrossRef] [PubMed] [Google Scholar]
- Marteil L. 1955. La reconstitution des gisements naturels d'huîtres plates. Rev Trav Inst Pêches Marit 19: 310–344. [Google Scholar]
- Marteil L. 1959. Les bancs naturels et la reproduction de l'huître plate en Morbihan. Rev Trav Inst Pêches Marit 23: 287–295. [Google Scholar]
- Marteil L. 1960. Écologie des huîtres du morbihan. Rev Trav Inst Pêches Marit 24: 335–445. [Google Scholar]
- Marteil L. 1976. La conchyliculture française, 2 partie: biologie de l'huître et de la moule. Chapitre VI: maladies et mortalites. Rev Trav Inst Pêche Marit 40: 285–313. [Google Scholar]
- Martin AG, Gérard A, Cochennec N, Langlade A. 1992. Selecting flat oysters, Ostrea edulis, for survival against the parasite Bonamia ostreae: assessment of the resistance of a first selected generation. In: Production, Environment and Quality, Bordeaux Aquaculture, '92. Edited by the European Aquaculture Society. Special Publication No. 18, Ghent Belgium, pp. 547-554. [Google Scholar]
- Martin AG. 1991. Relance de l'huître plate. Rapport de synthèse des travaux 1990. Rapport Ifremer. RIDRV-91.19-RA/LA TRINITE SUR MER. https://archimer.ifremer.fr/doc/00102/21320/ [Google Scholar]
- Martin AG. 1993. Relance de l'huître plate. Rapport d'avancement des travaux. Année 1992. Rapport Ifremer. RIDRV-93.031-RA. https://archimer.ifremer.fr/doc/00427/53823/ [Google Scholar]
- Martin AG. 1994. Relance de l'huître plate: Bilan des travaux réalisés en 1993. Rapport Ifremer. RIDRV- 94.22 RA. https://archimer.ifremer.fr/doc/00427/53822/ [Google Scholar]
- Marty GD, Bower SM, Clarke KR, Meyer G, Lowe G, Osborn AL, Chow EP, Hannah H, Byrne S, Sojonky K, Robinson JH. 2006. Histopathology and a real-time PCR assay for detection of Bonamia ostreae in Ostrea edulis cultured in western Canada. Aquaculture 261: 33–42. [CrossRef] [Google Scholar]
- Mérou N, Lecadet C, Pouvreau S, Arzul I . 2020. An eDNA/eRNA‐based approach to investigate the life cycle of non‐cultivable shellfish micro‐parasites: the case of Bonamia ostreae, a parasite of the European flat oyster Ostrea edulis. Microb Biotechnol 13: 1807-1818 https://doi.org/10.1111/ 1751-7915. 13617 [Google Scholar]
- Mérou N, Lecadet C, Billon T, Chollet B, Pouvreau S, Arzul I . 2022. Investigating the environmental survival of Marteilia refringens, a marine protozoan parasite of the flat oyster Ostrea edulis, through an environmental DNA and microscopy-based approach. Front Mar Sci 9: 811284 (12p.) [PubMed] [Google Scholar]
- Mérou N. 2021. Étude de l'écologie de Marteilia refringens et Bonamia ostreae, deux parasites protozoaires de l'huître plate, Ostrea edulis. PhD Thesis, Université de La Rochelle, 246 p. https://archimer.ifremer.fr/doc/00690/80166/ [Google Scholar]
- Mialhe E, Bachère E, Chagot D, Grizel H. 1988. Isolation and purification of the protozoan Bonamia ostreae (Pichot et al., 1980), a parasite affecting the flat oyster Ostrea edulis L. Aquaculture 71: 293–299. [CrossRef] [Google Scholar]
- Morga B, Arzul I, Choillet B, Renault T. 2009. Infection with the protozoan parasite Bonamia ostreae modifies in vitro haemocyte activities of flat oyster Ostrea edulis. Fish Shellfish Immunol 26: 836–842. [CrossRef] [PubMed] [Google Scholar]
- Morga B, Arzul I, Faury N, Segarra A, Chollet B, Renault T. 2011. Molecular responses of Ostrea edulis haemocytes to an in vitro infection with Bonamia ostreae. Dev Comp Immunol 35: 323–333. [CrossRef] [PubMed] [Google Scholar]
- Morga B, Renault T, Arzul I . 2012. New insights in flat oyster Ostrea edulis resistance against the parasite Bonamia ostreae. Fish Shellfish Immunol. 32: 958–968. [CrossRef] [PubMed] [Google Scholar]
- Morga B, Renault T, Faury N, Lerond S, Garcia C, Chollet B, Joly JP, Lapègue S, Harrang E, Arzul I . 2017. Contribution of in Vivo Experimental Challenges to Understanding Flat Oyster Ostrea edulis Resistance to Bonamia ostreae. Front Cell Infect Microbiol 7: 433. doi: 10.3389/fcimb.2017.00433 [CrossRef] [Google Scholar]
- Morga B. 2010. Etude des interactions hôte/parasite chez l'huître plate Ostrea edulis et son parasite Bonamia ostreae / Study of host/parasite interactions in the flat oyster Ostrea edulis and the parasite Bonamia ostreae. PhD Thesis, Université de La Rochelle., 400p. https://archimer.ifremer.fr/doc/00055/16664/ [Google Scholar]
- Mougne C, Dupont C. 2013. Les Gaulois et la pêche à pied en Plaine de Caen. Les Gaulois au fil de l'eau 1: 569–592. [Google Scholar]
- Naciri-Graven Y, Haure J, Gerard A, Baud JP. 1999. Comparative growth of Bonamia ostreae resistant and wild flat oyster Ostrea edulis in an intensive system. II. Second year of the experiment. Aquaculture 171: 195–208. [CrossRef] [Google Scholar]
- Naciri-Graven Y, Martin AG, Baud JP, Renault T, Gérard A. 1998. Selecting the flat oyster Ostrea edulis (L.) for survival when infected with the parasite Bonamia ostreae. J Exp Mar Biol Ecol. 224: 91–107. [CrossRef] [Google Scholar]
- Pardo BG, Álvarez-Dios JA, Cao A, Ramilo A, Gómez-Tato A, Planas JV, Villalba A, Martínez P. 2016. Construction of an Ostrea edulis database from genomic and expressed sequence tags (ESTs) obtained from Bonamia ostreae infected haemocytes: development of an immune-enriched oligo-microarray. Fish Shellfish Immunol. 59: 331–344. [CrossRef] [Google Scholar]
- Pascual M, Martin AG, Zampatti E, Coatanea D, Defossez J, Robert R. 1991. Testing of the Argentina oyster, Ostrea puelchana, in several French oyster farming sites. Acte du Conseil International pour l'Exploration de la Mer, C.M 1991/K: 30. [Google Scholar]
- Petton S, Pouvreau S, Dumas F. 2020. Intensive use of Lagrangian trajectories to quantify coastal area dispersion. Ocean Dyn 70: 541–559. [CrossRef] [Google Scholar]
- Pichot Y, Comps M, Tigé G, Grizel H, Rabouin MA. 1980. Recherches sur Bonamia ostreae gen. n., sp. n., parasite nouveau de l'huître plate Ostrea edulis L. Rev Trav Inst Pêche Marit 43: 131–140. [Google Scholar]
- Pineda J, Reyns NB, Starczak VR. 2009. Complexity and simplification in understanding recruitment in benthic populations. Popul Ecol 51: 17–32. [CrossRef] [Google Scholar]
- Pogoda B, Brown J, Hancock B, Preston J, Pouvreau S, Kamermans P, Sanderson W, von Nordheim H. 2019. The native oyster restoration alliance (NORA) and the Berlin oyster recommendation: bringing back a key ecosystem engineer by developing and supporting best practice in Europe. Aquat Living Resour 32: 13. [CrossRef] [EDP Sciences] [Google Scholar]
- Potet M, Fabien A, Chaudemanche S, Sebaibi N, Guillet T, Gachelin S, Cochet H, Boutouil M, Pouvreau S. 2021. Which concrete substrate suits you? Ostrea edulis larval preferences and implications for shellfish restoration in Europe. Ecol Eng 162: 106159. [CrossRef] [Google Scholar]
- Pouvreau S, Cochet H, Bargat F, Petton S, Le Roy V, Guillet T, Potet M. 2021b. Current distribution of the residual flat oysters beds (Ostrea edulis) along the west coast of France. SEANOE. https://doi.org/10.17882/79821 [Google Scholar]
- Pouvreau S, Cochet H, Gachelin S, Chaudemanche S, Fabien A. 2019. Inventaire, diagnostic écologique et restauration des principaux bancs d'huîtres plates en Bretagne : le projet FOREVER. Rapport Intermédiaire 2018. RBE/PFOM/PI 2019-3, 30 p. [Google Scholar]
- Pouvreau S, Cochet H, Gachelin S, Chaudemanche S, Fabien A. 2021a. Inventaire, diagnostic écologique et restauration des principaux bancs d ' huîtres plates en Bretagne: le projet FOREVER. Rapport final. Contrat FEAMP 17/2215675, 122 p. https:// 1453 doi.org/10.13155/79506 [Google Scholar]
- Pouvreau S, Huber M, Petton S, Le Roy V, Mingant C, Breton F, Sourget Q. 2016. DIversification de la pêcherie de la RAde de Brest par l'étude de semis de PEtoncle Noir: le projet DIRAPEN. Rapport Interne Ifremer, 30 p. https://archimer.ifremer.fr/doc/00363/47425/ [Google Scholar]
- Pouvreau S. Juillet E., Gilante H. 2021c. Projet FOREVER (Flat Oyster Recovery) : Restauration écologique de l'huître plate en Bretagne. Retours d'expériences. Génie écologique, Centre de ressources, 14 p. https://archimer.ifremer.fr/doc/00742/85402/ [Google Scholar]
- Prado P, Fernández M, Cordero D, Saavedra C, Carella F, Alcaraz C, Gairin I. 2022. Molecular identification, life cycle characterization, and hatchery seed production of dwarf oysters from the Ebro Delta (Spain). Aquat Living Resour 35: 5. [CrossRef] [EDP Sciences] [Google Scholar]
- Prado-Alvarez M, Chollet B, Couraleau Y, Morga B, Arzul I . 2013. Heat shock protein 90 of Bonamia ostreae: characterization and possible correlation with infection of the flat oyster, Ostrea edulis. J Eukaryot Microbiol 60: 257–266. [CrossRef] [PubMed] [Google Scholar]
- Prandtl, L. 1926 Ueber Die Ausgebildete Turbulenz. Proceedings 2nd International Congress Applied Mechanics, Zurich, 12–17 September 1926, pp. 62. [Google Scholar]
- Ramilo A, Navas JI, Villalba A, Abollo E. 2013. Species-specific diagnostic assays for Bonamia ostreae and B exitiosa in European flat oyster Ostrea edulis: conventional, real-time and multiplex PCR. Dis Aquat Org 104: 149–161. [CrossRef] [PubMed] [Google Scholar]
- Robert M, Garcia C, Chollet B, Lopez-Flores I, Ferrand S, Francois C, Joly JP, Arzul I. 2009. Molecular detection and quantification of the protozoan Bonamia ostreae in the flat oyster, Ostrea edulis. Mol Cell Probes 23: 264–271. [CrossRef] [PubMed] [Google Scholar]
- Robert R, Borel M, Pichot Y, Trut G. 1991. Growth and mortality of the European oyster Ostrea edulis in the Bay of Arcachon (France). Aquat Living Resour 4: 265–274. [CrossRef] [EDP Sciences] [Google Scholar]
- Robert R, His E, Dinet A. 1988. Combined effects of temperature and salinity on fed and starved larvae of the European flat oyster Ostrea edulis Mar Biol 97: 95–100. [CrossRef] [Google Scholar]
- Robert R, Pichot Y, Comps M. 1990. Essai de culture de l'huître plate Ostrea edulis dans le bassin d'Arcachon. Résultats préliminaires. IDRV-90.27-RA, 25 p. https://archimer.ifremer.fr/doc/00000/1592/ [Google Scholar]
- Robert R, Vignier J, Petton B. 2017. Influence of feeding regime and temperature on development and settlement of oyster Ostrea edulis (Linnaeus, 1758) larvae. Aquac Res 48: 4756–4773. [CrossRef] [Google Scholar]
- Ronza P, Cao A, Robledo D, Gómez-Tato A, Álvarez-Dios JA, Hasanuzzaman AFM, Quiroga MI, Villalba A, Pardo BG, Martínez P. 2018. Long-term affected flat oyster (Ostrea edulis) haemocytes show differential gene expression profiles from naïve oysters in response to Bonamia ostreae. Genomics 110: 390–398. [CrossRef] [PubMed] [Google Scholar]
- Saavedra C, Zapata C, Alvarez G. 1995. Geographical patterns of variability at allozyme loci in the European oyster Ostrea edulis. Mar Biol 122: 95–104. [CrossRef] [Google Scholar]
- Saavedra C, Zapata C, Guerra A, Alvarez G. 1993. Allozyme variation in European populations of the oyster Ostrea edulis. Mar Biol 115: 85–95. [CrossRef] [Google Scholar]
- Sambade IM, Casanova A, Blanco A, Gundappa MK, Bean TP, Macqueen DJ, Houston RD, Villalba A, Vera M, Kamermans P, Martínez P. 2022. A single genomic region involving a putative chromosome rearrangement in flat oyster (Ostrea edulis) is associated with differential host resilience to the parasite Bonamia ostreae. Evol Appl 15: 1408–1422. [CrossRef] [PubMed] [Google Scholar]
- Sierra R, Canas-Duarte SJ, Burki F, Schwelm A, Fogelqvist J, Dixelius C, Gonzalez-Garcia LN, Gile GH, Slamovits CH, Klopp C, Restrepo S, Arzul I, Pawlowski J. 2016. Evolutionary origins of Rhizarian parasites. Mol Biol Evol 33: 980–983. [CrossRef] [PubMed] [Google Scholar]
- Sobolewska H, Beaumont AR. 2005. Genetic variation at microsatellite loci in northern populations of the European flat oyster (Ostrea edulis). J Mar Biol Assoc UK 85: 955–960. [CrossRef] [Google Scholar]
- Stanisière JY, Cochet H, Cloatre T, Dumas F, Petton S, Pouvreau S. 2016. Étude de la variabilité du recrutement de l'huître plate en Baie de Quiberon : le projet EVER. Rapport final, 109 p. [Google Scholar]
- Suquet M, Gonzalez R, Lebrun L, Queau I, Mingant C, Robert R. 2010. Anaesthesia and gonad sampling in the European flat oyster (Ostrea edulis). Aquaculture 308: 196–198. [CrossRef] [Google Scholar]
- Taris N, Boudry P, Bonhomme F, Camara MD, Lapègue S 2009. Mitochondrial and nuclear DNA analysis of genetic heterogeneity in temporal recruitment cohorts of the European flat oyster, Ostrea edulis. Biol Bull 217: 223–241. [Google Scholar]
- Thébault A, Bergman S, Pouillot R, Le Roux F, Berthe FCJ. 2005. Validation of in situ hybridisation and histology assays for the detection of the oyster parasite Marteilia refringens. Dis Aquat Org 65: 9–16. [CrossRef] [PubMed] [Google Scholar]
- Tigé G, Grizel H, Cochennec N, Rabouin MA. 1984. Evolution de la situation épizootiologique en Bretagne en 1983 suite au développement de Bonamia ostreae. CIEM C.M 1984/F:14: 14 pp. [Google Scholar]
- Tigé G, Grizel H, Rabouin MA, Cochennec N, Audic G, Langlade A. 1982. Maladie hémocytaire de l'huître plate causée par Bonamia ostreae : évolution de la situation épizootiologique en Bretagne au cours de l'année 1981. Bull Inst Pêches Mar 328: 3–13. [Google Scholar]
- Vera M, Carlsson J, Carlsson JE, Cross T, Lynch S, Kamermans P, Villalba A, Culloty S, Martinez P. 2016. Current genetic status, temporal stability and structure of the remnant wild European flat oyster populations: conservation and restoring implications. Mar Biol 163: 239. [CrossRef] [Google Scholar]
- Vera M, Pardo BG, Cao A, Vilas R, Fernández C, Blanco A, Gutierrez AP, Bean TP, Houston RD, Villalba A, Martinez P. 2019. Signatures of selection for bonamiosis resistance in European flat oyster (Ostrea edulis): new genomic tools for breeding programs and management of natural resources. Evol Appl 12: 1781–1796. [CrossRef] [PubMed] [Google Scholar]
- zu Ermgassen PSE, Bonačić K, Boudry P, Bromley CA, Cameron TC, Colsoul B, Coolen JWP, Frankić A, Hancock B, van der Have TM, Holbrook Z, Kamermans P, Laugen AT, Nevejan N, Pogoda B, Pouvreau S, Preston J, Ranger CJ, Sanderson WG, Sas H, Åsa Strand, Sutherland WJ. 2020. Forty questions of importance to the policy and practice of native oyster reef restoration in Europe. Aquat Conserv 30: 2038–2049. [CrossRef] [Google Scholar]
- zu Ermgassen PSE, Bos O, Debney A, Celine G, Glover A, Pogoda B, Pouvreau S, Sanderson WG, Smyth D, Preston J 2021. European Native Oyster Habitat Restoration Monitoring Handbook. The Zoological Society of London Press, London, UK, 69p. https://archimer.ifremer.fr/doc/00745/85667/ [Google Scholar]
Cite this article as: Pouvreau S, Lapègue S, Arzul I, Boudry P. 2023. Fifty years of research to counter the decline of the European flat oyster (Ostrea edulis): a review of French achievements and prospects for the restoration of remaining beds and revival of aquaculture production. Aquat. Living Resour. 36: 13
All Tables
Non-exhaustive inventory of research and development work (published articles and reports) carried out in France since the emergence of the epizootics linked to the parasites Marteilia refringens and Bonamia ostreae. Most cited references are available via https://archimer.ifremer.fr/ or on request from the French technical centres CAPENA, SMIDAP or SMEL.
Citations related to the flat oyster fishery explicitly mentioning the physical degradation of the habitat.
All Figures
|
Fig. 1 Bonamia ostreae (arrows) in hemocytes of Ostrea edulis (A: histology; B: gill imprint). Marteilia refringens (arrows) in the epithelium of the digestive diverticula in Ostrea edulis (C: histology; D: digestive gland imprint). Pictures by Bruno Chollet / Ifremer. |
| In the text | |
|
Fig. 2 Occurrence of wild beds of flat oysters in Brittany, the colours indicate the stages of their conservation status. For more information, see Pouvreau et al. (2021b). |
| In the text | |
|
Fig. 3 Ecological and conservation stages of beds and reefs of O. edulis observed in Brittany, France. For more information, see Pouvreau et al. (2021b). |
| In the text | |
|
Fig. 4 Temporal variations in abundance of O. edulis larvae in the bays of Brest (Roz station) and Quiberon (Men Er Roué station). The relation with water temperature is indicated by the red strip. Adapted from Pouvreau et al. (2021a). |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.