| Issue |
Aquat. Living Resour.
Volume 34, 2021
|
|
|---|---|---|
| Article Number | 9 | |
| Number of page(s) | 11 | |
| DOI | https://doi.org/10.1051/alr/2021007 | |
| Published online | 09 April 2021 | |
Research Article
Changes in age and maturity of anadromous whitefish (Coregonus lavaretus) in the northern Baltic Sea from 1998 to 2014
1
Natural Resources Institute (Luke), Wolffintie 35, Vaasa FI-65200, Finland
2
Natural Resources Institute (Luke), Helsinki FI-00791, Finland
3
Department of Biology, University of Turku, Turku FI-20014, Finland
4
Natural Resources Institute (Luke), Laivurintie 6, Keminmaa FI-94450, Finland
* Corresponding author: This email address is being protected from spambots. You need JavaScript enabled to view it.
Handling Editor: AE Richard Nash
Received:
27
July
2020
Accepted:
7
March
2021
Abstract
The maturation of anadromous whitefish (Coregonus lavaretus) was analysed from samples taken from commercial coastal fishing in 1998–2014 in the Gulf of Bothnia. Whitefish matured at a younger age from year to year. The proportion of older (5–12 sea years) mature males decreased from 79% to 39% in the northern Gulf of Bothnia (66°N–64°N) and from 76% to 14% in southern (64°N–60°30'N) during the study period. At the same time, the proportion of young males (2–4 sea years) increased. Whitefish matured younger: the proportion of mature fish at age four increased in both the north and south among females (13% → 98%; 6% → 85%) and males (68% → 99%; 29% → 89%). The catch length of four-year-old fish increased during the study period in both sexes. In contrast, the length of six-year-old females decreased from year to year. Sea surface temperatures increased during the study period, and were possibly associated with a decrease in the age of maturation and faster growth.
Key words: Anadromous whitefish / maturation / temperature / Baltic Sea
© EDP Sciences 2021
1 Introduction
The maturation and growth rate of anadromous whitefish (Coregonus lavaretus) varies between local stocks in Finnish coastal areas (Lehtonen, 1981; Heikinheimo and Mikkola, 2004; Aronsuu and Huhmarniemi, 2004). This variation may either be due to an evolutionary adaptation to local conditions (Säisä et al., 2008; Olsson et al., 2012; Hägerstrand et al., 2017) or partly or entirely due to phenotypic plasticity (Price et al., 2003). The age structure of the spawning stock may be wide, rangingin fish from 2 to 12 years of age (Lehtonen, 1981; Czerniejewski and Rybczyk, 2010).
Fishing of European whitefish has changed considerably during last seven decades in the Finnish coastal waters in the Baltic Sea. The most important fishing gears in the 1950s were trap nets in the Gulf of Bothnia. Caught whitefish weighted 1–3 kg, and mesh sizes in trap nets and gillnets were usually between 50 and 65 mm (Lehtonen and Jokikokko, 2002). Pelagic drift net fishing of migrating anadromous whitefish was started in the 1960s. In the 1990s the mesh size in gill nets has decreased, being 40–55 and 30–35 mm in the southern and northern part of Gulf of Bothnia, respectively and harvested whitefish weighted 0.4–0.8 kg (Lehtonen and Jokikokko, 2002). The mesh size of gill nets in commercial whitefish fishing in 1998–2011 was mainly 40–45 and 25–39 mm in the southern and northern Gulf of Bothnia, respectively (Kallio-Nyberg et al., 2018).The average length-at-size of old anadromous whitefish entering to their home rivers in the Gulf of Bothnia has decreased since 1980s (Lehtonen and Jokikokko, 2002) and mean weight of mature anadromous whitefish in the River Tornionjoki has had a decreasing trend from 1980 to 2019 (Jokikokko and Huhmarniemi, 2014; Jokikokko et al., 2020).
Whitefish stocks have undergone large-scale changes since the 1950s in Finnish coastal waters (Urho, 2011). Many original stocks have been lost and natural production has decreased (Kaukoranta et al., 2000), and the sea catch has decreased since the 1990s to nearly fourth part of top level (Urho, 2011; OSF, 2020). The commercial whitefish catch in Finnish coastal waters in 2017 was 476 tonnes, and 81% of the catch was taken in the Gulf of Bothnia (GoB) (OSF, 2020). The loss of natural production has been compensated for fisheries with large-scale stockings in Finland (Jokikokko et al., 2002; Jokikokko and Huhmarniemi, 2014). The level of stockings was highest in the 1980s when over 15 million fingerlings were released to the coastal area (Salojärvi, 1986). Recently, the fingerling stockings have decreased substantially and are approx. 7 million fingerlings annually (ICES, 2018). Also newly hatched larvae are stocked, and the quantities have decreased from level of 40–90 million larvae in 1990s (Jokikokko et al., 2002) to approx. 30 million annually (ICES, 2018). Most of the releases are done with anadromous whitefish fingerlings or larvae in rivers running to the northern Gulf of Bothnia (Jokikokko and Huhmarniemi, 2014).Whitefish larvae or fingerlings disperse to feeding migration immediately after hatching or release (Lehtonen et al., 1992), and return to home river as mature fish (Kallio-Nyberg et al., 2019). Stockings likely have a strong impact on spawning stock and catches (Leskelä et al., 2004; Jokikokko and Huhmarniemi, 2014).
Gillnet fishing has been shown to affect the life history traits of exploited stocks (Lehtonen and Jokikokko, 2002; Nusslé et al., 2009; Uusi-Heikkilä et al., 2015). In commercial fishingin the Gulf of Bothnia approx. 80% of whitefish are caught with gillnets and the rest with trapnets (OSF, 2020). Gill-net fishing effectively removes the largest individuals (Heikinheimo and Mikkola, 2004) and puts anadromous whitefish under selective fishing (Lehtonen and Jokikokko, 2002; Aronsuu and Huhmarniemi, 2004). It has e.g. been observed that the length of the six-year-old anadromous females has decreased in the commercial catches of the Gulf of Bothnia in 1998–2014 (Kallio-Nyberg et al., 2019). The decreased size and age at maturation in an intensive harvested pikeperch (Sander lucioperca) stock in the southern Gulf of Bothnia was likely a consequence of the selective gill net fishing (Kokkonen et al., 2015).
Temperature is known to play a significant role in the growth rate, survival, year-class strength, distribution and maturation of fish species (Jonsson and Jonsson, 2009; Pankhurst and Munday, 2011; Audzijonyte et al., 2020) and effect of annual environmental temperature variation on life-history traits of fish species can be seen also in the Baltic Sea (Jutila et al., 2005; Kallio-Nyberg et al., 2006; Pekcan-Hekim et al., 2011; Heikinheimo et al., 2014). Several documents suggest that the observed body-size reduction is a universal response to global warming (Gardner et al., 2011). Many fish species has shifted their distribution northward with climate warming (Perry et al., 2005). Global temperature has increased, and most scenarios concerning the Baltic Sea area predict an increase in the sea temperature and a shorter period of ice cover in winter (Meier et al., 2004; Mackenzie et al., 2007; Räisänen, 2017).
The aim of this study was to analyse the maturation and growth trends of anadromous whitefish during a time period when the sea temperature increasedand fishing was concentrated more and more on larger anadromous whitefish instead of local sea spawning whitefish, partly due to collapse of southern sea spawning whitefish stocks (Veneranta et al., 2013). This smaller whitefish was in earlier years a target for intensive fishing thus balancing the effort towards anadromous form (Jokikokko et al., 2018; Kallio-Nyberg et al., 2020). Special attention is placed on the effect covariation of climate data such as the North Atlantic Oscillation (NAO) and the local sea surface temperature (SST) with the age of maturation.
2 Material and methods
2.1 Whitefish samples
Whitefish samples were collected as part of the EU Data Collection Framework (DCF) by Natural Resources Institute Finland. The DCF whitefish sample is an unbiased sample of the Finnish commercial whitefish fishery from 1998 to 2014. The samples used in this and earlier studies (Kallio-Nyberg et al., 2018, 2019) represent commercial whitefish fishing in the GoB both spatially and temporally. Samples were from fish quarterly bought from unselected catch of fishermen along the coast of the GoB and covering all fishing methods. For the analyses the GoB was split into the northern part (the northern Bothnian Bay), covering ICES rectangles 1–16 (between 66°N and 64°N), and the southern part (the southern Bothnian Bay and the Bothnian Sea), covering ICES rectangles 17–47 (64°N–60°30'N; Fig. 1). The northern and southern samples were analysed separately, because the fishing and growth rate of anadromous whitefish differ between these areas (Lehtonen, 1981; Hägerstrand et al., 2017; Kallio-Nyberg et al., 2019). The median mesh size used in the gill net fishing for all whitefish forms is smaller (28 mm from knot to knot) in the northern than in the southern GoB (40 mm) in the unbiased sample from the commercial catch in the Gulf of Bothnia in 1998–2011 (Kallio-Nyberg et al., 2018). The southern anadromous stocks migrate only in the southern part of Gulf of Bothnia, but northern stocks migrate in the both, southern and northern sea areas (Leskelä et al., 2002; Leinonen et al., 2020).
In the Finnish coastal area, two forms of whitefish (Coregonus lavaretus) occur, namely anadromous and sea-spawning forms (Kallio-Nyberg et al., 2019). These two forms were separated from each other by gill raker counts, growth rate, size and maturation and spawning place (Lehtonen, 1981; Himberg et al., 2015). The mean gill-raker number for anadromous whitefish was 29.6 (n = 11799, SD = 2.1, min = 21, max = 43). From anadromous whitefish 33% and 67% were caught in the northern and southern GoB, respectively. Mature whitefish totalled 4944 and 6021, respectively, at the age of 2–12 years. Their sex was also determined (Tab. 1).
The whitefish stocks from the rivers of the Bothnian Bay migrate to feed even as far as to the Bothnian Sea in the south (Leskelä et al., 2002), but a proportion of northern anadromous whitefish remain in the Bothnian Bay near their home rivers (Lehtonen and Jokikokko, 2002; Hägerstrand et al., 2017; Jokikokko et al., 2018). For most whitefish the gillrakers were counted and the age, sex, maturation, total length (mm) and weight (g) were determined. Ageing of fish was based on otoliths which were cut and grinded to show the growth rings and the reading was made with a microscope. Based on the maturation state, the anadromous whitefish were divided into immature fish with no sign of maturation in the ovarian, and mature fish, which were identified as being able to spawn in the catch year according to Kesteven (1960). The first sea year for whitefish was considered to be the next whole year after the year of hatching or release (1–12 months). The whitefish were aged and their form and maturation was identified by professional personnel in LUKE.
The fishermen reported the fishing site, date and the gear used. The whitefish samples were caught with trap nets (34%) and gillnets (66%). The most important fishing months were August (18.9%) and September (24.8%) in the southern GoB and June (19.4%) and September (33.8%) in the northern GoB, when all male and female whitefish were included in the study (south: n = 11 578; north: n = 6045).
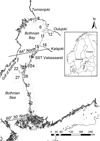 |
Fig. 1 The northern Gulf of Bothnia, covering ICES rectangles 1–16 (between latitudes 66°N and 64°N), and the southern Gulf of Bothnia, covering ICES rectangles 17–47 (between latitudes 64°N and 60°30'N). The whitefish (Coregonus lavaretus) samples were collected in the indicated rectangles in 1998–2014. Rivers: 1 = Tornionjoki, 2 = Oulujoki, 3 = Kalajoki. Sea surface temperature measurement site: Valassaaret station. |
Age distribution (%) and median age (md) with quartiles (q1–q3) and mean age of mature anadromous male (M) and female (F) whitefish (Coregonus lavaretus) in the northern and southern Gulf of Bothnia in 1998–2014.
2.2 Statistics
The age distributions of mature female and male anadromous whitefish caught in both the northern and southern GoB were calculated and the differences between sexes and areas were tested using the χ2 test during 1998–2014. Age groups were excluded from the χ2 test if their sample size was under 5. The median age for the sexes in the south and north was calculated and the differences were tested using the Kruskal-Wallis test. Also the mean age for sexes was calculated and the normality of age distribution was tested using Kolmogorov-Smirnov −test andnon-parametric testing was applied due to the lack of normality.The younger age groups (2, 3, 4, 5, 6, 7 years) were separate and older (8–12 years) age groups were pooled in age distribution during 1998–2014, and the proportion of the mature fishin these seven groups were calculated separately. All mature fish in the sample independent from the catch months were included.
The change in the mean catch age with time (1998–2014) was analysed using linear regression, and the sexes were analysed separately for the northern and southern GoB (Results in Appendix: Mean age of whitefish in relation to catch year; Fig. A.1).
Changes in the proportions of the young (2–4 years, n = 4620) and old (5–12 years, n = 6345) age groups were explained by year (1998–2014, continuous variable), sex, and area (northern and southern GoB) in a binomial regression model as applied in Saloniemi et al. (2004). The two-year-old fish were the youngest fish in the sample, and halving the data to groups 2–4 and 5–12 years old gave close to equal number of fish in both age groups and four- and five-year-old fish were the most common the data (Appendix: Proportion of old mature fish and maturation rate at age group, Tab. A.1).
Trends of maturation age by year (1998–2014) were analysed by calculating the proportion of mature individuals in 4- and 5-year-old fish in the northern and southern GoB. The probability of anadromous whitefish being mature (binomial mature vs. immature) at the age of four or five years was analysed according to binomial regression models, when sex, sea area (southern and northern) and year, and their interactions were used as predictors (Appendix: Proportion of old mature fish and maturation rate at age group, Tab. A.2). Only fish caught in June–December were included, because whitefish spawn in late autumn (Lehtonen, 1981).
The increase in length was analysed as a function of age (in months) and sex for both study periods. At first, the sample was divided to two periods: early (1996–2002) and later (2003–2009), because we wanted to study first the change separately in both periods. To account for non-linear trends, the time spent at sea was also included in the squared form (month*month) in the models. The individual fish aged 3–6 sea years (25–72 months) caught in the GoB (rectangles 17–47) were included, and the body length of whitefish was log-transformed to normality (Appendix: Growth of anadromous whitefish, Tab. A.3). Next the increase in size was also analysed by including the period as a separate predictor in the previous model (Appendix: Growth of anadromous whitefish, Tab. A.4).
Associations between the mean age of mature fish of year class and environmental factors experienced by this year class was analysed using Spearman correlation. The annual environmental factors were the mean sea surface temperature (SST) in June measured weekly (4–5 times per month)in Valassaaret (63°44'N, 21°07'E) and the seasonal NAO index in June to August (Hurrell and National Center for Atmospheric Research Staff, 2016). The year class experienced the annual SST average in June in third sea year (25–36 months at sea). Linear regression was also used to test if the June SST (in the third summer of the year class) and NAO index (in the second summer of the year class) predicted the age of the mature females of the year class.
Valassaaret station is located in the middle of the feeding areas of several anadromous whitefish stocks (Koljonen et al., 2019; Leinonen et al., 2020). SST has increased in the Baltic Sea in last decades (Jylhä et al., 2009). During positive seasonal NAO index westerly winds and a mild marine climate are dominant in summer, while during negative seasonal NAO index, east winds and continental climate dominates in Europe (Hurrell and National Center for Atmospheric Research Staff, 2016). Here it was used NAO index in June to August, which the year class was experienced in their second sea year (13–24 months at sea; January–December) The NAO index in the summer months is used in this research, because the growth of whitefish is highest in summer. NAO index in the early sea-migration period, in the second summer of the year class was used, since the maturation of the year-class begins at this point.All statistical analyses were conducted with SAS 9.4 software package (SAS Institute Inc, Cary, North Carolina).
3 Results
During the study period, 1998–2014, most mature whitefish were four or five years old in the GoB (Tab. 1). Fish were younger in the south compared to the north (Kruskal-Wallis; median length: males: df = 5, χ2 = 544.9, P < 0.001; females: df = 5, χ2 = 468.9.1, P < 0.001 [Tab. 1]). The youngest age in the catch was two both in the north and south, and even some of them were mature (see Tab. 2).
The share of older whitefish (5–12 sea years) among the mature stock decreased from year to year in 1998–2014,whereas the proportion of younger fish (2–4 sea years) increased in the sea catch in the southern and northern Gulf of Bothnia (Fig. 2). The proportion of older (5–12 sea years) mature females in the stock near the home rivers in north decreased from 97% to 60% and that of males from 79% to 39% from the beginning of the study period in 1998 until the end in 2014 (Fig. 2; models shown in Tab. A.1).
At the same time, the proportion of mature fish among young age groups increased. The proportion of mature whitefish at the age of four increased among fish caught in June–December in the GoB. The proportion of mature four-year-old females increased from 13% to 98% and that of males from 68% to 99% in the northern sea area from year to year in 1998–2014, i.e., practically all four-year-old fish were mature at the end of study period (sex: P < 0.001; catch year: P < 0.001; model: Appendix: Tab. A.2 [Fig. 3]). In the southern GoB, the proportion of mature fish increased more among females (6% → 85%) than males (29% → 89%), with a significant difference between the sexes (sex*catch year: P < 0.001). The same trend is seen among five-year-old fish (Fig. 3).
Growth was modelled using age (25–72 months) and sex as predictors (Appendix Tab. A.3) and finally by including the year period as a separate predictor in the previous models (Fig. 4; Appendix Tab. A.4). The catch length of sexes was different in both year-class periods, the difference between sexes was larger in the later period (Fig. 4; Tab. A.4). The young fish grew faster in the period 2003–2009 than in the period 1996–2002. In contrast, the growth of old fish declined in the later period (Tab. A.5). The period was statistically significant when it was included in the model (Tab. A.4).
There was a significant negative correlation between the mean age of mature whitefish of the year class and the SST (Valassaaret station, June) experienced by this year class in its third sea summer (Fig. 5; Tab. 3). In addition, there was a significant positive correlation between the mean age of mature whitefish of the year class and the seasonal NAO index in June to August in the second summer of the year class (Tab. 3), meaning that the marine climate in summer months (NAO high) was associated with the higher age of mature anadromous whitefish.
The mean sea surface temperature in June in the third sea year of the year class was used to predict the sea age of the mature females of the year class in the southern Gulf of Bothnia (Linear regression model: Age (year) = 7.77–0.280 × SST, F(1,12) = 23.11, P < 0.001, R2 = 0.658 [Fig. 6]). High summer SST was associated with a younger mean sea age of the year class. If the NAO index was added to the same model too, only SST explained the sea age of females of the year class.
The proportion of mature anadromous whitefish (%; Coregonus lavaretus) in different sea age groups amongmales (M) and females (F) in the northern and southern Gulf of Bothnia in 1998–2014.
 |
Fig. 2 The predicted share of 5–12-year-old mature female and male anadromous whitefish (Coregonus lavaretus) in the catch in the northern (N) and southern (S) Gulf of Bothnia (GoB) in relation to the catch year (1998–2014). The response variable was classified as young (age 2–4) or older (age 5–12 years) fish. The age group, sex and area are predictors (model Tab. A.1). Sample sizes for young and old fish 4620 and 6345 in GoB, respectively. |
 |
Fig. 3 The predicted proportion of mature female and male anadromous whitefish (Coregonus lavaretus) in relation to the catch year (1998–2014) among four- (age 4) and five-year-old (age 5) mature and immature fish caught in June–December in the southern (S) and northern (N) Gulf of Bothnia (models, Tab. A.2). Sample sizes for mature and immature fish 3479 and 1668 at age 4 and 3260 and 644 at age 5, respectively. |
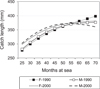 |
Fig. 4 Predicted length of individual anadromous female (F) and male (M) whitefish (Coregonus lavaretus) in relation to time (months) spent at sea in two year-class periods (1996–2002, 2003–2009; symbols: 1990 and 2000, respectively) in the Gulf of Bothnia according to linear regression model. The whitefish captured by trap and gill nets in the third-sixth sea year (25–72 months; 25–36 months = the third year; 37–48 = the fourth year, 49–60 = the fifth year, 61–72 = the sixth year) are included (sample size = 12653). The length is log-transformed (natural) in the model, but transformed back to linearity in the figures. Sample size for earlier and later periods, n = 6151 and n = 6516, respectively (Tab. A.4). |
 |
Fig. 5 The mean age of anadromous female whitefish (Coregonus lavaretus) in the southern (Age_S) and northern (Age_N) Gulf of Bothnia in the year classes 1992–2006 in their third sea year and sea surface temperature (SST) at the Valassaaret station. The linear trends of the mean age and SST are shown. Correlation between the mean age and SST: north: P = 0.003; south: P = 0.004. |
Spearman correlation coefficient (r) between the mean age of anadromous Coregonus lavaretus year class and the annual sea surface temperature (SST) or between the seasonal (June–August) NAO index in the northern and southern Gulf of Bothnia.
 |
Fig. 6 Model for the dependence of the mean age of mature whitefish on sea surface temperature (SST) in June in Valassaaret. The model predicts a linear trend (continuous line) between the mean age of mature females of the year class and June SST in the third summer of the year class. The 95% confidence limits of the mean (broken lines) and observations (diamond) are presented. |
4 Discussion
The age of mature fish, maturation at a specific age, and the growth rate of the anadromous whitefish changed dramatically in the GoB during 1998–2014, and these age changes were likely linked to climatic changes and fishing.The used data was temporally and spatially unbiased sample from the GoB whitefish fishing (Kallio-Nyberg et al., 2018, 2019), thus the decreased proportion of the old (6+ and older) mature whitefish in the northern GoB, near the home rivers of the anadromous whitefish stocks suggest that the age distribution of spawning stocks have changed. A decreasing trend in the whitefish catch size during upstream migration has been observed in the Kalajoki and Tornionjoki rivers (Aronsuu and Huhmarniemi, 2004; Jokikokko and Huhmarniemi, 2014), suggesting also that spawners are younger and smaller than earlier. In this study, the sea age of mature anadromous whitefish decreased with time, and in the same period sea surface temperature increased in the GoB. Increased environmental temperature has reported to be an indirect or direct influence on developmental rate and maturation (Jonsson and Jonsson, 2009; Pankhurst and Munday, 2011). The climatic conditions and temperature experienced by fish in their development affect their later life history phase (Jonsson et al., 2005) and even traits in next generation (Jonsson and Jonsson, 2016). Here, it was used only June SST in the third summer in the analysis between temperature and age of mature females, but is likely that temperature has increased also in other phases of the year class over time. Fish species have optimum temperature and thermal limits for growth and increase of warming within lowest thermal limit and optimum temperature usually increases growth rate (Elliot and Elliot, 2010) and when growth rate is in interaction with maturation; then climatic-induced increased growth usually tends to lead to earlier maturation (Neuheimer and Grønkjær, 2012).
However, the selective effect of gill-net fishing on the age of mature fish cannot be excluded. Selective fishing has been shown to be the most important driving factor favouring early maturation (Jørgensen et al., 2009), for instance, in pikeperch (Kokkonen et al., 2015). Earlier studies on the anadromous whitefish in the GoB have provided evidence of both selective fishing (Lehtonen and Jokikokko, 2002; Heikinheimo and Mikkola, 2004) and annual and long-term changes in the sea water temperature (Mackenzie et al., 2007; Siegel and Gerth, 2017). Kallio-Nyberg et al. (2019) reported a link between the improved growth of young and small whitefish and increasing air temperature in the GoB. This same whitefish catch data shows that the growth of the fast-growing anadromous whitefish and slow-growing sea-spawning whitefish differed between 1998 and 2014 in the GoB. The catch size-at-age of the small sea-spawning whitefish was increasing, but the large anadromous whitefish was decreasing, which suggests selective fishing against fast growth rate, large size of spawners and late maturation (Kallio-Nyberg et al., 2019). Also here the growth rate of old anadromous whitefish decreased from year class period 1996–2002 to period 2003–2009 (Fig. 4). Age and age-specific size has decreased also in the spawning stock of the anadromous whitefish in the River Kemijoki (Kallio-Nyberg et al., 2020).
On average, males reached sexual maturity one year earlier than females among all mature whitefish in this study (1998–2014). During 1999–2002, mature females were mainly aged five and six and males were aged four and five in the Bothnian Bay (Leskelä et al., 2004). The shift in the maturation rate differed between the sexes: the proportion of maturating 4–5 year-old whitefish increased faster among females than males, and the size of six-year-old females decreased, which suggests that the present selection pressure is stronger in the life-history traits of females compared to males. The female fecundity and egg size increase with body size (Szczepkowski et al., 2010). Thus, an earlier allocation of resources to reproduction instead of growth reduces the proportion of larger and older females in the spawning stock, which probably decreases the fitness of the stock (Birkeland and Dayton, 2005). Regarding the whitefish, the decreasing size of female spawners, which has already happened (Kallio-Nyberg et al., 2020), likely leads to decreasing reproductive capacity of the natural stocks.
The gill net is a selective item of fishing gear that effectively takes the fastest growing, largest individuals, thus affecting the size and sea-age distribution of the stock (Heikinheimo and Mikkola, 2004). The most anadromous whitefish in GoB are caught by selective gill nets (Jokikokko et al., 2020), and e.g., in commercial fishing approx. 80% from catch is from gillnet fishing and 20% from fyke-net fishing. The mesh size in gill nets decreased in the Bothnian Bay during 1998–2011 (Kallio-Nyberg et al., 2018), which likely increased selective effect of this gear. It is probable that this has targeted fishing on relatively late-maturing, large and old females in the GoB, and small individuals have had a higher probability of reproducing than late-maturing fish. Known responses to gill-net fishing include earlier maturation (Kokkonen et al., 2015) and a decreased growth in older whitefish (Heikinheimo and Mikkola, 2004; Aronsuu and Huhmarniemi, 2004; Kallio-Nyberg et al., 2019). The age structure and maturity stages observed here in whitefish are likely to be responses to the combined effects of fishing and climate-induced changes. Selective fishing increases the proportion of small and likely young fish in the spawning stock (see Jokikokko et al., 2018), while a warming climate increases the growth rate and leads to maturation at a younger age. When the size-at-age increases, the fish are recruited at a younger age for fishing.
Simultaneous selective fishing and increasing temperatures may compensate each other. In many cases, the effects of fishing-induced changes may lead to the under estimation of the effect of global warming. An increasing SST increases the growth rate, independent of the time spent at sea, but this fast growth rate is not realised in older age groups due to the counteracting effects of selective fishing. However, new data from Koljonen et al. (2019) and Leinonen et al. (2020) indicates that whitefish fishing targets mixed anadromous whitefish stocks. Thus, the change in the vitality of stocks and variation in stocking numbers may also interact with observed changes in the growth pattern. Stockings are done annually and mainly with anadromous whitefish stocks for compensatory purposes in dammed rivers running to the northern Gulf of Bothnia but not anymore for common-good as in 1960s–1980s (Jokikokko and Huhmarniemi, 2014).
The changes in fish species size and maturation due to fishing or climate-induced evolution can have major ecological, genetic and economic consequences (Daufresne et al., 2009; Sheridan and Bickford, 2011; Uusi-Heikkilä et al., 2015). Larger fish species, in particular, decrease in abundance and size (Todd et al., 2008; Genner et al., 2010). The whitefish catches in commercial fishing have decreased during this century in the GoB, but part of this decrease is probably due to changes in fishing and a reduction in stocking (Jokikokko and Huhmarniemi, 2014). Other factors, such as the low price and decreasing demand for small sea spawning whitefish, increasing number of seals that both consume whitefish and disturb fishing (Hansson et al., 2017; Tverin et al., 2019), or fishing regulations, have reduced the profitability and hence the intensity of gill-net fishing (Söderlind, 2004; Jokikokko et al., 2018).
Acknowledgements
Thanks to the staff of the Natural Resources Institute Finland (Luke), especially Hannu Harjunpää and Alpo Huhmarniemi for their substantial contribution to the whitefish data collection, and the Finnish Data Collection team for its considerable work in collecting whitefish data according to the EU Data Collection Framework in Finland. The present study was partly funded by the SmartSea project, funded by the Strategic Research Council of the Academy of Finland, and partly by Luke.
Appendix: A
A.1 Mean age of whitefish in relation to catch year
The annual mean age of male and female whitefish decreased over the years (1998–2014) both in the northern and southern Gulf of Bothnia (GoB; Fig. A.1). The annual mean age of female and male fish covaried in both areas (northern GoB: r = 0.784, n = 17, P < 0.001, and southern GoB: r = 0.836, n = 17, P < 0.001).
 |
Fig. A.1 The annual mean age of anadromous male and female whitefish (Coregonus lavaretus) age classes in the catch years 1998–2014 in the northern (north) and southern (south) Gulf of Bothnia. The mean annual age decreased over 1998–2014 (regression for males in north: mean age = 122.382 − 0.058 × year, F 1,15 = 32.23, P < 0.001, r 2 = 0.661; females in north: mean age = 64.824 − 0.029 × year, F 1,15 = 11.01, P = 0.005, r 2 = 0.423; males in south: mean age = 84.170 − 0.039 × year, F 1,15 = 10.22, P = 0.006, r 2 = 0.405; and females in south: mean age = 80.801–0.038 × year, F 1,15 = 5.59, P = 0.032, r 2 = 0.272). |
A.2 Fishing season and sample time
The fishing season within the period from June–December started earlier in the northern GoB (month = 33.051 − 0.012 × catch year; F 1,1569 = 2.50, P = 0.001) and in the southern GoB (month = 36.127 − 0.014 × year; F 1,2331 = 4.12, P = 0.001) when five-year-old fish were included. The median sample month in the northern GoB for five-year-old males was 9 (= September; quartiles: 8–9) (n = 917) and that for females was 9 (7–9; n = 654). In the southern GoB, this was 8 both for males and for females (n = 1281, n = 1052, respectively) when June–December sample sizes were included in the models.
A.3 Proportion of old mature fish and maturation rate at age group
Predicted share of the 5–12-year-old mature whitefish in the northern and southern Gulf of Bothnia in relation to the catch year and sex.
Model for the proportion of mature four- and five-year-old whitefish in the Gulf of Bothnia during 1998–2014.
A.4 Growth of anadromous whitefish
Growth models for 3-6-year-old whitefish in the GoB separately in two year-class periods.
Growth models for 3–6-year-old whitefish in the GoB.
Age-specific median (Md) weight (g) of anadromous male and female whitefish in two class periods (1996–2002; 2003–2009) in the Gulf of Bothnia.
References
- Aronsuu K, Huhmarniemi A. 2004. Changes in the European whitefish (Coregonus lavaretus (L.)) population of the Kalajoki—potential consequences of the alterations of fishing patterns in the Gulf of Bothnia. Ann Zool Fennici 41: 195–204. [Google Scholar]
- Audzijonyte A, Richards SA, Stuart-Smith RD, Pecl G, Edgar GJ, Barrett NS, Payne N, Blanchard JL. 2020. Fish body sizes change with temperature but not all species shrink with warming. Nat Ecol Evol 4: 809–814. [PubMed] [Google Scholar]
- Birkeland C, Dayton PK. 2005. The importance in fishery management of leaving the big ones. Trends Ecol Evol 20: 356–358. [PubMed] [Google Scholar]
- Czerniejewski P, Rybczyk A. 2010. Growth rate and condition of population migratory common whitefish, (Coregonus lavaretus L.), from Oder estuary waters. Arch Pol Fish 18: 25–32. [Google Scholar]
- Daufresne M, Lengfellner K, Sommer U. 2009. Global warming benefits the small in aquatic ecosystems. PNAS 106: 12788–12793 [Google Scholar]
- Elliot JM, Elliot JA. 2010. Temperature requirements of Atlantic salmon Salmo salar, brown trout Salmo trutta and Arctic charr Salvelinus alpinus: predicting the effects of climate change. J Fish Biol 77: 1793–1817. [PubMed] [Google Scholar]
- Gardner JL, Peters A, Kearney MR, Joseph L, Heinsohn R. 2011. Declining body size: a third universal response to warming? Trends Ecol Evol 26: 285–291. [PubMed] [Google Scholar]
- Genner MJ, Sims DW, Southward AJ, Budd GC, Masterson P, McHugh M, Rendle P, Southall EJ, Wearmouth VJ, Hawkins SJ. 2010. Body size-dependent responses of a marine fish assemblage to climate change and fishing over a century-long scale. Global Change Biol 16: 517–527. [Google Scholar]
- Hägerstrand H, Heimbrand Y, von Numers M, Lill J-O, Jokikokko E, Huhmarniemi A. 2017. Whole otolith elemental analysis reveals feeding migration patterns causing growth rate differences in anadromous whitefish from the Baltic Sea. Ecol Freshw Fish 26: 456–461. [Google Scholar]
- Hansson S, Bergström U, Bonsdorff E, Härkönen T, Jepsen N, Kautsky L, Lundström K. 2017. Competition for the fish − fish extraction from the Baltic Sea by humans, aquatic mammals, and birds. ICES J Mar Sci 75: 999–1008. [Google Scholar]
- Heikinheimo O, Mikkola J. 2004. Effect of selective gill-net fishing on the length distribution of European whitefish (Coregonus lavaretus) in the Gulf of Finland. Ann Zool Fennici 41: 357–366. [Google Scholar]
- Heikinheimo O, Pekcan-Hekim Z, Raitaniemi J. 2014. Spawning stock-recruitment relationship in pikeperch Sander lucioperca (L.) in the Baltic Sea, with temperature as an environmental effect. Fish Res 155: 1–9. [Google Scholar]
- Himberg M, von Numers M, Vasemägi A, Heselius SJ, Wiklund T, Lill JO, Hägerstrand H. 2015. Gill raker counting for approximating the ratio of river and seaspawning whitefish, Coregonus lavaretus (Actinopterygii: Salmoniformes: Salmonidae) in the Gulf ofBothnia, Baltic Sea. ActaIchthyologicaEtPiscatoria 45: 125–131. [Google Scholar]
- Hurrell J, National Center for Atmospheric Research Staff (Eds.) 2016. Last modified 16 Aug. 2016. The climate Data Guide. Hurrell North Atlantic Oscillation (NAO) Index (Station-based). Retrieved from https://cliamtedataguide.ucar.edu/climate-data-hurrell-north-atlantic-oscillation-nao-index-station-based. [Google Scholar]
- ICES. 2018. Interim Report of the Working Group on Introductions and Transfers of Marine Organisms (WGITMO), Madeira, Portugal, 7–9 March 2018, ICES CM 2018/HAPISG:11, pp. 179. [Google Scholar]
- Jokikokko E, Huhmarniemi A. 2014. The large-scale stocking of young anadromous whitefish (Coregonus lavaretus) and corresponding catches of returning spawners in the River Tornionjoki, northern Baltic Sea. Fish Manag Ecol 21: 250–258. [Google Scholar]
- Jokikokko E, Leskelä A, Huhmarniemi A. 2002. The effect of stocking size on the first winter survival of whitefish, Coregonus lavaretus (L.), in the Gulf of Bothnia, Baltic Sea. Fish Manag Ecol 9: 79–85. [Google Scholar]
- Jokikokko E, Hägerstrand H, Lill J-O. 2018. Short feeding migration associated with a lower mean size of whitefish in the River Tornionjoki, northern Baltic Sea. Fish Manag Ecol 2018: 261–266. [Google Scholar]
- Jokikokko E, Veneranta L, Huhmarniemi A. 2020. Pohjanlahden siika, in: J. Raitaniemi, K. Manninen (Eds.), Kalakantojen tila 2019 ja ennuste vuosille 2020 ja 2021, Luonnonvara- jabiotaloudentutkimus 46/2020, Luonnonvarakeskus, Helsinki, pp. 50–58. [Google Scholar]
- Jonsson B, Jonsson N. 2009. A review of the likely effects of climate change on anadromous Atlantic salmon Salmo salar and brown trout Salmo trutta, with particular reference to water temperature and flow. J Fish Biol 75: 2381–2447. [PubMed] [Google Scholar]
- Jonsson B, Jonsson N. 2016. Trans-generational maternal effect: temperature influences egg size of the offspring in Atlantic salmon Salmo salar. J Fish Biol doi: 10.111/jfb.13040. [PubMed] [Google Scholar]
- Jonsson B, Jonsson N, Hansen, LP. 2005. Does climate during embryonic development influence parr growth and age of seaward migration in Atlantic salmon (Salmo salar)? Can J Fish Aqaut Sci 62: 2502–2508. [Google Scholar]
- Jørgensen C, Ernande B, Fiksen Ø. 2009. Size-selective fishing gear and life history evolution in the Northeast Arctic cod. Evol Appl 2: 356–370. [PubMed] [Google Scholar]
- Jutila E, Jokikokko E, Julkunen K. 2005. The smolt run and postsmolt survival of Atlantic salmon, Salmo salar L., in relation to early summer water temperatures in the northern Baltic. Ecol Freshw Fish 14: 69–78. [Google Scholar]
- Jylhä K, Ruosteenoja K, Räisänen K, Venäläinen A, Tuomenvirta H, Ruokolainen L, Seitola S. 2009. Arvioita Suomen muuttuvasta ilmastosta sopeutumistutkimuksia varten. ACCLIM-hankkeenraportti 2009. Ilmatieteen Laitoksen Raportteja, 4: 102 [In Finnish] [Google Scholar]
- Kallio-Nyberg I, Jutila E, Jokikokko E, Saloniemi I. 2006. Survival of reared Atlantic salmon and sea trout in relation to marine conditions of smolt year in the Baltic Sea. Fish Res 80: 295–304. [Google Scholar]
- Kallio-Nyberg I, Saloniemi I, Veneranta L, Salminen M. 2018. Anadromous trout threatened by whitefish gill-net fisheries in the northern Baltic Sea. J Appl Ichthyol 34: 1145–1151. [Google Scholar]
- Kallio-Nyberg I, Veneranta L, Saloniemi I, Jokikokko E. 2019. Different growth trends of whitefish (Coregonus lavaretus) forms in the northern Baltic Sea. J Appl Ichthyol 35: 683–691. [Google Scholar]
- Kallio-Nyberg I, Veneranta L, Jokikokko E, Leskelä A. 2020. Vaellussiian pituus- ja ikäjakauma Pohjanlahden saaliissa 1981–2017 sekä 2013 alkaneen verkkokalastussäätelyn vaikutus siikakantoihin. Luonnonvara- ja biotalouden tutkimus 95/2020. Luonnonvarakeskus, Helsinki, 44 pp. ISBN 978-952-380-109-7 [In Finnish]. [Google Scholar]
- Kaukoranta M, Koljonen M-L, Koskiniemi J, Pennanen J, Tammi J. 2000. Atlas of Finnish fishes, English summary. Distribution of lamprey, brook lamprey, salmon trout, Arctic charr, whitefish, vendace, grayling, asp, vimba, spined loach and bullhead, and status of the socks. Finnish Game and Fisheries Research Institute. Research Report. 40 p. ISBN: 951-776-287-9 [Google Scholar]
- Kesteven GL. 1960. Manual of field methods in fisheries biology. FAO Man Fish Sci 1: 44–45. [Google Scholar]
- Kokkonen E, Vainikka A, Heikinheimo O. 2015. Probabilistic maturation reaction norm trends reveal decreased size and age at maturation in an intensively harvested stock of pikeperch Sander luciaperca. Fish Res 167; 1–12. [Google Scholar]
- Koljonen M.L, Veneranta L, Kallio-Nyberg I, Koskiniemi J, Jokikokko E. 2019. Pohjanlahden siikakantojen erilaistuminen ja merialueen siikasaaliiden alkuperä. Luonnonvara- ja biotalouden tutkimus −sarja. Luonnonvara- ja biotalouden tutkimus 56/ 2019, 52 pp [In Finnish] [Google Scholar]
- Lehtonen H. 1981. Biology and stock assessments of Coregonids by the Baltic coast of Finland. Finnish Fish Res 3: 31–83. [Google Scholar]
- Lehtonen H, Jokikokko E. 2002. Responses of anadromous European whitefish, Coregonus lavaretus (L.) to fishing in the Gulf of Bothnia. ArchivFürHydrobiologie. Special Issues. Adv Limnol 57: 669–676. [Google Scholar]
- Lehtonen H, Nyberg K, Vuorinen PJ, Leskelä A. 1992. Radioactive strontium (85Sr) in marking whitefish [Coregonus lavaretus (L.)] larvae and the dispersal of larvae from river to sea. J Fish Biol 41: 417–423. [Google Scholar]
- Leinonen T, Kallio-Nyberg I, Koljonen M-L, Veneranta L, Jokikokko, E. 2020. Pohjanlahden siikakantojen vaelluserot ja ikäluokkien kokoerot: Siikakantojen ekologisten ominaisuuksien tutkimus geneettisen kannantunnistuksen avulla. Luonnonvara- ja biotalouden tutkimus 51/2020, Luonnonvarakeskus. Helsinki, 32 s. [Google Scholar]
- Leskelä A, Jokikokko E, Huhmarniemi A. 2002. Sea migration patterns of stocked anadromous European whitefish (Coregonus lavaretus L.) fingerlings. Arch Hydrobiol Spec Issues Adv Limnol 57: 119–128. [Google Scholar]
- Leskelä A, Jokikokko E, Huhmarniemi A, Siira A, Savolainen H. 2004. Stocking results of spray-marked one-summer old anadromous European whitefish in the Gulf of Bothnia. Ann Zool Fenn 41: 171–179. [Google Scholar]
- Mackenzie BR, Gislan H, Möllmann C, Köster FW. 2007. Impact of 21st century climate change on the Baltic Sea fish community and fisheries. Global Change Biol 13: 1348–1367. [Google Scholar]
- Meier MHE, Döscher R, Halkka A. 2004. Simulated distributions of Baltic Sea-ice in warming climate and consequences for the winter habitat of the Baltic Ringed Seal. AMBIO 33: 249–256. [PubMed] [Google Scholar]
- Neuheimer AP, Grønkjær P. 2012. Climate effects on size-at-age: growth in warming waters compensates for earlier maturity in an exploited marine fish. Global Change Biol 18: 1812–1822. [Google Scholar]
- Nusslé S, Bornand CN, Wedekind C. 2009. Fishery-induced selection on an Alpine whitefish: quantifying genetic and environmental effects on individual growth rate. Evol Appl 2: 200–208. [PubMed] [Google Scholar]
- Olsson J, Florin A-B, Mo K, Aho T, Ryman N. 2012. Genetic structure of whitefish (Coregonusmaraena) in the Baltic Sea. Estuar Coast Shelf S 97: 104–113. [Google Scholar]
- OSF. 2020. Official Statistics Finland. Commercial Marine Fishery. Natural Resources Institute Finland, Helsinki. https://statdb.luke.fi/PXWeb/pxweb/fi/LUKE/LUKE__06%20Kala%20ja%20riista__02%20Rakenne%20ja%20tuotanto__02%20Kaupallinen%20kalastus%20merella/4_meri_saalis.px/ (accessed December 16, 2020) [Google Scholar]
- Pankhurst NW, Munday PL. 2011. Effect of climate change on fish reproduction and early life history stages. Mar Freshw Res 62: 1015 [Google Scholar]
- Pekcan-Hekim Z, Urho L, Auvinen H, Heikinheimo O, Lappalainen J, Raitaniemi J, Söderkultalahti P. 2011. Climate warming and pikeperch year-class catches in the Baltic Sea. Ambio 40: 447–456. [PubMed] [Google Scholar]
- Perry AL, Low PJ, Ellis JR, Reynolds JD. 2005. Climate change and distribution shifts in marine fishes. Science 308: 1912–1915. [Google Scholar]
- Price TD, Qvarnström A, Irwin DE. 2003. The role of phenotypic plasticity in driving genetic evolution. Proc R Soc London B 270: 1433–1440. [Google Scholar]
- Räisänen J. 2017. Future Climate Change in the Baltic Sea Region and Environmental Impacts. Oxford Research Encyclopedia of Climate Science. https://doi.org/10.1093/acrefore79780190228620.013.634. [Google Scholar]
- Salojärvi K. 1986. Review of whitefish (Coregonus lavaretus L. s.l.) fingerling rearing and stocking in Finland. Arch Hydrobiol Beih Ergebn Limnol 22: 99–114. [Google Scholar]
- Saloniemi I, Jokikokko E, Kallio-Nyberg I, Jutila E, Pasanen P. 2004. Survival of reared and wild Atlantic salmon smolts: size matters more in bad years. ICES Journal of Marine Science 61: 782–787. [Google Scholar]
- Säisä M, Rönn J, Aho T, Björklund M, Pasanen P, Koljonen M-L. 2008. Genetic differentiation among European whitefish ecotypes based on microsatellite data. Hereditas 145: 69–83. [PubMed] [Google Scholar]
- Sheridan JA, Bickford D. 2011. Shrinking body size as an ecological response to climate change. Nat Clim Change 1: 401–406 [Google Scholar]
- Siegel H, Gerth M. 2017. Sea surface temperature in the Baltic Sea in 2016. Baltic Sea Environment Fact Sheet 2017, Published on 12 October 2017. http://helcom.fi/baltic-sea-trends/environment-fact-sheets/hydrography/development-of-sea-surface-temperature-in-the-baltic-sea [Google Scholar]
- Szczepkowski M, Szczepkowska N, Krywosz T, Wunderlich K, Stabiński R. 2010. Growth rate and reproduction of a brood stock of European whitefish (Coregonus lavaretus L.) from Lake Gaładuś under controlled rearing conditions. Arch Pol Fish 18: 3–11. [Google Scholar]
- Söderlind A. 2004. Estimation of the seal-inflicted hidden damage in the net fishery for pike-perch and whitefish, Master thesis in Marine Zoology, Department of Marine Ecology, Göteborg University, pp. 13. [Google Scholar]
- Todd CD, Hughes SL, Marshall T, MacLean JC, Lonergan ME, Biow EM, 2008. Detrimental effects of recent ocean surface warming on growth condition of Atlantic salmon. Global Change Biol 14: 1–13. [Google Scholar]
- Tverin M, Esparza-Salas R, Strömberg A, Tang P, Kokkonen I, Herrero A, Lundström K. 2019. Complementary methods assessing short and long-term prey of a marine top predator–Application to the grey seal-fishery conflict in the Baltic Sea. PloS one 14: e0208694. [PubMed] [Google Scholar]
- Urho L. 2011. Kalasto-, kalakantamuutokset ja vieraslajit ilmaston muuttuessa. RKTL: ntyöraportteja 6/2011. [In Finnish] Assess method: www.rktl.fi/www/uploads/pdf/uudet%20julkaisut/tyoraportit/kalasto_ilmastomuutos. [Google Scholar]
- Uusi-Heikkilä S, Whiteley AR, Kuparinen A, Matsumura S, Venturelli PA, Wolter C, Slate J, Primmer CR, Meinelt T, Killen SS, Bierbach D, Polverino G, Ludvig A, Arlinghaus R. 2015. The evolutionary legacy of size-selective harvesting extends from genes to populations. Evol Appl 8: 597–620. [PubMed] [Google Scholar]
- Veneranta L, Hudd R, Vanhatalo J. 2013. Reproduction areas of sea-spawning coregonids reflect the environment in shallow coastal waters. Mar Ecol Prog Ser 477: 231–250. [Google Scholar]
Cite this article as: Veneranta L, Kallio-Nyberg I, Saloniemi I, Jokikokko E. 2021. Changes in age and maturity of anadromous whitefish (Coregonus lavaretus) in the northern Baltic Sea from 1998 to 2014. Aquat. Living Resour. 34: 9
All Tables
Age distribution (%) and median age (md) with quartiles (q1–q3) and mean age of mature anadromous male (M) and female (F) whitefish (Coregonus lavaretus) in the northern and southern Gulf of Bothnia in 1998–2014.
The proportion of mature anadromous whitefish (%; Coregonus lavaretus) in different sea age groups amongmales (M) and females (F) in the northern and southern Gulf of Bothnia in 1998–2014.
Spearman correlation coefficient (r) between the mean age of anadromous Coregonus lavaretus year class and the annual sea surface temperature (SST) or between the seasonal (June–August) NAO index in the northern and southern Gulf of Bothnia.
Predicted share of the 5–12-year-old mature whitefish in the northern and southern Gulf of Bothnia in relation to the catch year and sex.
Model for the proportion of mature four- and five-year-old whitefish in the Gulf of Bothnia during 1998–2014.
Growth models for 3-6-year-old whitefish in the GoB separately in two year-class periods.
Age-specific median (Md) weight (g) of anadromous male and female whitefish in two class periods (1996–2002; 2003–2009) in the Gulf of Bothnia.
All Figures
|
Fig. 1 The northern Gulf of Bothnia, covering ICES rectangles 1–16 (between latitudes 66°N and 64°N), and the southern Gulf of Bothnia, covering ICES rectangles 17–47 (between latitudes 64°N and 60°30'N). The whitefish (Coregonus lavaretus) samples were collected in the indicated rectangles in 1998–2014. Rivers: 1 = Tornionjoki, 2 = Oulujoki, 3 = Kalajoki. Sea surface temperature measurement site: Valassaaret station. |
| In the text | |
|
Fig. 2 The predicted share of 5–12-year-old mature female and male anadromous whitefish (Coregonus lavaretus) in the catch in the northern (N) and southern (S) Gulf of Bothnia (GoB) in relation to the catch year (1998–2014). The response variable was classified as young (age 2–4) or older (age 5–12 years) fish. The age group, sex and area are predictors (model Tab. A.1). Sample sizes for young and old fish 4620 and 6345 in GoB, respectively. |
| In the text | |
|
Fig. 3 The predicted proportion of mature female and male anadromous whitefish (Coregonus lavaretus) in relation to the catch year (1998–2014) among four- (age 4) and five-year-old (age 5) mature and immature fish caught in June–December in the southern (S) and northern (N) Gulf of Bothnia (models, Tab. A.2). Sample sizes for mature and immature fish 3479 and 1668 at age 4 and 3260 and 644 at age 5, respectively. |
| In the text | |
|
Fig. 4 Predicted length of individual anadromous female (F) and male (M) whitefish (Coregonus lavaretus) in relation to time (months) spent at sea in two year-class periods (1996–2002, 2003–2009; symbols: 1990 and 2000, respectively) in the Gulf of Bothnia according to linear regression model. The whitefish captured by trap and gill nets in the third-sixth sea year (25–72 months; 25–36 months = the third year; 37–48 = the fourth year, 49–60 = the fifth year, 61–72 = the sixth year) are included (sample size = 12653). The length is log-transformed (natural) in the model, but transformed back to linearity in the figures. Sample size for earlier and later periods, n = 6151 and n = 6516, respectively (Tab. A.4). |
| In the text | |
|
Fig. 5 The mean age of anadromous female whitefish (Coregonus lavaretus) in the southern (Age_S) and northern (Age_N) Gulf of Bothnia in the year classes 1992–2006 in their third sea year and sea surface temperature (SST) at the Valassaaret station. The linear trends of the mean age and SST are shown. Correlation between the mean age and SST: north: P = 0.003; south: P = 0.004. |
| In the text | |
|
Fig. 6 Model for the dependence of the mean age of mature whitefish on sea surface temperature (SST) in June in Valassaaret. The model predicts a linear trend (continuous line) between the mean age of mature females of the year class and June SST in the third summer of the year class. The 95% confidence limits of the mean (broken lines) and observations (diamond) are presented. |
| In the text | |
|
Fig. A.1 The annual mean age of anadromous male and female whitefish (Coregonus lavaretus) age classes in the catch years 1998–2014 in the northern (north) and southern (south) Gulf of Bothnia. The mean annual age decreased over 1998–2014 (regression for males in north: mean age = 122.382 − 0.058 × year, F 1,15 = 32.23, P < 0.001, r 2 = 0.661; females in north: mean age = 64.824 − 0.029 × year, F 1,15 = 11.01, P = 0.005, r 2 = 0.423; males in south: mean age = 84.170 − 0.039 × year, F 1,15 = 10.22, P = 0.006, r 2 = 0.405; and females in south: mean age = 80.801–0.038 × year, F 1,15 = 5.59, P = 0.032, r 2 = 0.272). |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.