| Issue |
Aquat. Living Resour.
Volume 34, 2021
Special Issue - Ecological intensification: A new paragon for sustainable aquaculture
|
|
|---|---|---|
| Article Number | 5 | |
| Number of page(s) | 11 | |
| DOI | https://doi.org/10.1051/alr/2021002 | |
| Published online | 23 March 2021 | |
Research Article
Organic matter reduction using four densities of seaweed (Gracilaria verucosa) and green mussel (Perna viridis) to improve water quality for aquaculture in Java, Indonesia
1
Department of Aquaculture Faculty of Fisheries and Marine Sciences, Diponegoro University, Semarang, Indonesia
2
Aquaculture and Fisheries Group (AFI), Wageningen University, Wageningen, The Netherlands
* Corresponding author: This email address is being protected from spambots. You need JavaScript enabled to view it.
Handling Editor: Joel Aubin
Received:
25
May
2020
Accepted:
9
February
2021
Abstract
The high organic waste content of river water in Demak, north coast of Java, has caused traditional small-scale pond farmers to stop stocking shrimp. This paper examines whether seaweed and mussel will improve the quality of water these farmers use. The effect of Gracilaria verucosa and Perna viridis on the water quality was assessed by measuring the removal rates (RRs) of total organic material (TOM), total ammonia nitrogen (TAN), nitrite, and nitrate. The specific growth rates (SGRs) of seaweed and mussel were also measured. Thirty-six semi-outdoor tanks containing 800 L of brackish water and 7 cm substrate were randomly assigned to four replications of four densities of G. verucosa: 50 (S50), 100 (S100), 150 (S150), and 200 (S200) g m−2, and of P. viridis: 60 (M60), 90 (M90), 120 (M120), and 150 (M150) g m−2. Weekly, the TOM, TAN, nitrite, and nitrate contents were measured, seaweed and mussel weighted; RRs and SGRs were calculated at the end of the study. The effect of densities on the RRs was significant for both seaweed and mussel. P. viridis was more effective in reducing TOM (by 38%) than G. verucosa (7%); G. verucosa achieved higher RRs for TAN, nitrite, and nitrate. At S200, TOM and TAN decreased by 7.4% and 67%, respectively. At M90, TOM and TAN, decreased by 38% and 49%, respectively. However, nitrite increased significantly at S200 and M150. The SGR of seaweed was significantly lower at S200 than that at S150, S100, and S50. The best performing densities were S100 and M90.
Key words: Macroalgae / mussel / organic-matter / ammonia / nitrate / nitrite
© EDP Sciences 2021
1 Introduction
In Central Java Province, Indonesia, with 29 districts, Demak regency ranks third in milkfish (11 474 tons/yr) and shrimp (783 tons/yr) production from brackish water ponds (Statistic of Central Java Province, 2020). The current yield is decreasing due to increasing water pollution from the waste generated by industries, households, and aquaculture. In addition, due to lack of capital and technology, traditional farmers do not have reservoir ponds to maintain water quality before culturing shrimp and fish. The advised maximum concentration of organic matter in the water for shrimp culture is less than 55 mgL−1 (MMAF, 2013), but in Demak the initial concentrations in the inlet water are higher: 234–734 (Rahmaningsih, 2012 ) and 96–124 mg L−1 (Yuniarsih et al., 2014). To reduce organic waste in the water for shrimp culture, we intend to use ecological approaches, that is, organisms of lower trophic levels, to absorb excess nutrients. An ecological approach to maintain good water quality that has been studied in recent years is the use of macro-algae and mussel to eliminate nitrogen and ammonium (Neori et al., 2004; Rabiei et al., 2014; Susilowati et al., 2014; Pena-Rodríguez et al., 2017). Culturing a combination of species of different trophic levels has emerged as a sustainable way to establish a more environment-friendly aquaculture system (Neori et al., 2004). Macroalgae, such as seaweeds, are the primary producers representing the lowest trophic level; while extractive filter-feeding organisms, such as green mussels, represent the second level. While building animal protein, low trophic species prevent the energy losses during trophic transfers. They are ideal species that can exploit available resources in coastal waters (Filgueira et al., 2019).
For their growth, seaweeds, such as G. verucosa, absorb nutrients, for example, N as ammonia or nitrate from the dissolved organic waste in the water column (Jones et al., 2001). Considering its role in improving water quality, seaweed has been cultured together with shrimp, tilapia and bivalves (Golez et al., 2002; Tendencia et al., 2006; Pandjara et al., 2010; Aliah, 2012). Meanwhile, filter-feeding organisms like the bivalve green mussel (P. viridis) feed on organic components in suspension, plankton and bacteria (Tendencia, 2007; Tantanasarit and Babel, 2014). Simultaneously, they decrease dissolved inorganic nitrogen and biological oxygen demand in an intensive shrimp farm's wastewater (Chaiyakum and Tanwilai, 1992; Haamer, 1996). Blue mussel has been used in the Bay of Fundy to partially biomitigate fish wastes.
As most organisms release nutrients as part of their excretion (Van Khoi and Fotedar, 2012), increased activity in the sediment and respiration by the algae and plankton cause the amount of dissolved oxygen (DO) to decrease, adversely affecting organisms (Christensen et al., 2003; Srisunot and Babel, 2015). Combining G. verucosa and P. viridis may optimize the removal of OM and other nutrients from water, but their optimal densities for the local conditions remain unknown. These densities must take account of competition for oxygen, particularly at night, when low DO contents impede the nitrification process, thus producing nitrite which at high concentrations is toxic for shrimp. In addition, too much seaweed may result in low availability of natural feed for the mussels and/or shrimps.
This study was a preliminary research for a pilot of an integrated multi-trophic shrimp aquaculture system; in this study we observed separately the potential role of two species: G. verucosa and P. viridis. Our study aimed to identify the optimal densities of seaweed and green mussel for improving the quality of water available for farmers in order to restore and maintain shrimp culture in areas such as Java suffering from industrial and urban water pollution. We compared and analyzed the effect of different densities of G. verucosa and P. viridis on their capacity to reduce the levels of total organic matter (TOM), total ammonia nitrogen (TAN), nitrite (NO2), and nitrate (NO3) in the water prior to shrimp culture. The growth rates of G. verucosa and P. viridis were recorded because of their financial contribution to the future system. Below we explain the methodology, and discuss and interpret the results we obtained.
2 Materials and methods
2.1 Data collection
2.1.1 Seaweed, green mussel, and mesocosm
The G. verucosa and P. viridis for the experiment were collected from farmers in the local village. In selecting G. verucosa, we considered three qualities of the thallus: color, size, and appearance; in selecting P. viridis, we considered the shell length (3 ± 0.2 cm) and the body weight (2.2 ± 0.25 g). For G. verucosa, we set up 16 tanks (4 treatments, 4 replications) with four different densities per treatment: 50, 100, 150, and 200 gm−2. Another 16 tanks (4 treatments, 4 replications) were simultaneously set up with P. viridis at four densities: 60, 90, 120, and 150 gm−2. Densities were based on Pandjara et al. (2010) and Aliah (2012). G. verucosa and P. viridis clusters, each initially weighing 50 and 30 g, respectively, were placed in nets attached to ropes and suspended in the water for 30 days (Fig. 1). In addition, for control, there was a tank with no seaweed or green mussels. The seaweed treatments will henceforth be referred to as S50, S100, S150, and S200; the green mussel treatments will be referred to as M60, M90, M120, and M150. The experimental set-up was laid out in a completely randomized design with four replicates for each treatment of seaweed and green mussel.
We created separate non-connected mesocosm for each replication by implementing the treatments in 36 fiberglass tanks of 1 × 1 × 1 m; each tank contained 10 cm mud-clay substrate and 80 cm of water, resulting in a volume of 800 L. Brackish water at 25–29 ppm salinity was pumped into the tank from a canal 200 m away; 10% of the water was changed every week to ensure the input of organic waste into the system, and to mimic the real condition in the locally practiced pond system. This canal received waste from upstream activities, both industrial and urban, and its water was used by local shrimp and milkfish farmers who have water inlets along that canal.
Every week, all seaweed and mussels were removed from each tank for growth observation. Seaweed was weighed using an A&D® HL-100 digital scale with 0.01 g accuracy; in green mussel, shell length was measured using a Krisbow® KW0600351 digital caliper with 0.01 cm accuracy.
 |
Fig. 1 G. verucosa (a) and P. viridis (b) were cultured by the hanging method. |
2.1.2 Water quality parameters
The pH and salinity were measured daily (9 am and 9 pm) by using a digital pH meter (HANNA® H198129; 0.01accuracy) and as alinometer (digital ATAGO® PAL-06S; 1 ppt accuracy), respectively. The effect of photosynthesis and respiration of seaweed and green mussel was recorded by measuring dissolved oxygen (DO) twice daily (9 am and 9 pm) with YSI® Pro 20 (accuracy 0.5 ppm). Fluctuations in water temperature were measured thrice daily (7 am, 2 pm and 9 pm) by using a digital thermometer with 0.1 °C accuracy.
TOM, TAN, NO3, and NO2 concentrations of 300 ml samples from each tank were analyzed weekly; samples were covered with ice, labeled, and kept in the cold box to prevent any changes during transport and storage. TAN, NO3, and NO2values were measured by using a Spectrophotometer (Optima 3000), following methods described by the National Standardization Agency of Indonesia: 06-6989.30-2005, 6989.99-2011 and 06-6989.9-2004 (SNI, 2004, 2011). TAN was quantified by using the salicylate method and 650 nm wavelength; NO3 by cadmium reduction method and 500 nm wavelength and NO2 by the diazotization method and 507 nm wavelength. TOM was measured by using potassium permanganate (KMnO4) to oxidize and quantify the OM (Kutty, 1987; SNI, 2004). Phytoplankton samples were collected by taking 10 L water, filtered out using plankton net with mesh size 25 μ, then preserved by adding 1% formalin solution and kept in cool conditions until laboratory analysis. Phytoplankton were counted by using the sweeping method in Sedgewick Rafter glass (APHA, 2012). Additionally, the transparency of the water column was observed by using a Secchi disk. The values and the changes in water color were also recorded.
2.2 Calculations
We defined organic waste by the concentrations of TOM, TAN, NO3, and NO2. The weekly concentrations were plotted in graphs that showed the trends and compared the treatments. Removal rate, phytoplankton abundance, specific growth rate and survival rate calculated using equations (1)–(4) respectively. Removal rates of TOM, TAN, NO3, and NO2 were defined as the differences before and after the cultivation period and were calculated by using the following equation (Srisunot and Babel, 2015). (1)
(1)
RR: Removal Rate, Ct: Final concentration (g L−1), Ci: Initial concentration (g L−1).
The abundance of phytoplankton was calculated with the following equation (APHA, 2012): (2)where N = phytoplankton abundance (ind ml−1); Oi = cover glass area (mm2); Op = view area (mm2); Vr = filtered water volume (ml); Vo = observed water volume (ml); n = number of phytoplankton in the entire view area; p = number of view areas.
(2)where N = phytoplankton abundance (ind ml−1); Oi = cover glass area (mm2); Op = view area (mm2); Vr = filtered water volume (ml); Vo = observed water volume (ml); n = number of phytoplankton in the entire view area; p = number of view areas.
The SGR of the seaweed was determined by the formula (Busacker et al., 1990): (3)where Wt = final weight, Wo = initial weight and T = cultivation days.
(3)where Wt = final weight, Wo = initial weight and T = cultivation days.
Survival rate of green mussel was calculated with the formula: (4)where SR = survival rate, Nt = the number of mussels at the end of the study, No = the initial number of mussels.
(4)where SR = survival rate, Nt = the number of mussels at the end of the study, No = the initial number of mussels.
2.3 Statistical analysis
The mean values of TOM, TAN, NO3, and NO2 were calculated weekly during the experimental period, using SPSS®. The homogeneity of data was tested with Levene's test for the two-way ANOVA and the Test of Sphericity for the repeated measures ANOVA. Two-way ANOVA was used to ascertain if the means of the two independent variables (time and density) were different, and if there was a significant interaction between them. To take account of the repeated weekly measures of the same variable taken on the same subjects at different time periods, we used the repeated measures analyses. In addition, in case of significant interaction between the effect of the independent variables on the dependent variables, a multiple comparison post-hoc test, that is, Duncan test was used to assess the effect of the independent variables.
3 Results
The Levene's and Sphericity tests both showed that the data were homogeneous (p > 0.05) and could be compared. For all four parameters of organic waste concentration (TOM, TAN, NO3, and NO2), the two-way ANOVA revealed that time and density both had a significant effect and that their interactions were significant as well. The repeated measures ANOVA confirmed the interaction between the two factors (day and density) on these four parameters of organic waste concentration (p < 0.01). Further analyses showed that the concentrations of all parameters were significantly different from those in the control (Tab. 1).
Results of the two-way ANOVA testing the influence of days, density, and the interaction on TOM, TAN, NO2, NO3, and SGR of G. verucosa and P. viridis.
3.1 Total organic matter (TOM)
At the end of the experiment, the TOM in all treatments was significantly lower than that in the control (Tab. 1). The treatments were grouped pairwise: S50 and S100 (p = 0.63) and S150 and S200 (p = 0.69); TOM was significantly lower for the second pair (p < 0.01). The post-hoc test confirmed significant differences at 28 days of cultivation; prior to that time, the TOM values in each treatment were not significantly different (Tab. 1).
During the experiment, the average concentration of TOM (mg L−1) in the macro algae (S150 and S200) decreased from 265 to 234, while for S.50 and S.100 TOM's value remained stable (Fig. 2a). G. verocosa, at highest density rate (S200), removed the most TOM (7.4%); while lower densities of this microalgae removed less. TOM was highest in the lowest density (S50) of G. verocosa (Fig. 3a).
The effect of the interaction of time and density on the TOM content was clearly seen in tanks with green mussel (Fig. 2b). TOM in both the control and M150 had increased from 250 to above 350 mg L−1; at M120, TOM remained stable at 250 mg L−1 during the first 3 weeks, but increased slightly in the last week. However, at M90, TOM increased for two weeks; it peaked to above 350 mg L−1, but then declined and ended to below 170 mg L−1. At M60, the lowest density of green mussels, TOM decreased gradually, also ending at about 170 mg L−1. At both M60 and M90, the final values of TOM were significantly lower than those of the control, M120 and M150 (Fig. 2b). M90 had the highest TOM removal rate (38%) followed by M60 (Fig. 3b). In contrast, at the highest density of P. viridis, the content of TOM became even higher than that of the control (31% compared to 18%).
 |
Fig. 2 Concentration dynamics of TOM (mg L−1) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
 |
Fig. 3 The removal rates, averages and error bars of the four replications, of TOM (%) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
3.2 Total ammonia nitrogen (TAN)
In G. verucosa treatments, the TAN removal was significantly affected by the density (Tab. 1); in all densities, the concentration of TAN became lower than that in the control (p < 0.01). The continuous decrease of the TAN concentrations at the three higher densities contrasted with the increase at the lowest density in the last week of the study. In that week, the TAN concentration increased from 0.08 to 0.12 in the lowest density of seaweed, but in the other treatments it decreased to 0.06 mg L−1 (Fig. 4a). G. verucosa reduced the TAN concentration between 20% and 67%; the highest TAN removal (67%) was in tanks with the highest density S200 (Fig. 5a).
At M60 and M90, P. viridis reduced the TAN concentration significantly (p < 0.01) from 0.2 to 0.09 mg L−1 within 3 weeks. At the higher densities (M120 and M150), the TAN concentration remained at a level similar to that in the control. Although not significantly different, the TAN concentrations in M60 and M90 increased in the last week of the experiment (Fig. 4b). P. viridis reduced the TAN by up to 50% in the second lowest densities (M60 and M90), but removal rates were negative at the two highest densities (Fig. 5b).
 |
Fig. 4 Concentration dynamics of TAN (mg L−1) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
 |
Fig. 5 The removal rates, averages and error bars of the four replications, of TAN (%) in the control and at four densities of both G. verucosa (a) and P. viridis (b). |
3.3 Nitrite (NO2)
G. verucosa significantly reduced (p < 0.01) the nitrite in the water (Tab. 1) for all three lower densities: from 0.006 to 0.004 mg L−1 for S50, from 0.006 to 0.003 mg L−1 for S100, and from 0.007 to 0.003 mg L−1 for S150 (Fig. 6a). However, the concentration of nitrite in the highest density of G. verucosa, S200, was higher than that in the control. G. verucosa's nitrite removal rate was high: >50% for the two low concentrations (Fig. 7a).
We observed the same effect (p < 0.01) for the lower densities of P. viridis (M60 and M90) in comparison with the higher densities (M120 and M150) (Fig. 6b). In the first week, the concentration of nitrite increased in all treatments from 0.005 to 0.007 mg L−1, but thereafter dropped to 0.004 mg L−1 in the lower densities of P. viridis. The nitrite levels in the higher densities became as high as that in the control (0.008 mg L−1). The removal rates of P. viridis were below 10% (M-120 and M-60) or below zero (Fig. 7b). At the higher densities, the nitrite concentration rose in the last week of the experiment.
 |
Fig. 6 Concentration dynamics of NO2 (mg L−1) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
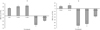 |
Fig. 7 The removal rates, averages and error bars of the four replications, of NO2 (%) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
3.4 Nitrate (NO3)
During the second half of the experimental period, the nitrate reduction tended to be greater in tanks with higher densities of seaweeds. In the fourth week, the nitrate concentrations in all treatments had decreased from about 1.7 to around 1.25 mg L−1. After 5 weeks, the nitrate concentration in S50 had remained at 1.25 mg L−1, while in S200 it had significantly decreased to below 1 mg L−1 (Fig. 8a). The nitrate concentration decreased in time as well as with increasing density; all nitrate values were lower than that of the control. At S200 G. verucosa had the highest nitrate removal rate: 52% (Fig. 9a). At S100, G. verucosa had already attained a higher removal rate than any of those achieved by P. viridis (Fig. 9a and 9b).
The final nitrate concentrations (mg L−1) in tanks with P. viridis (Fig. 8b) fell into three significantly different groups (p < 0.01): control (1.7 mg L−1), M60 and M.120 (1.5 mg L−1), M90 and M120 (1.1 mg L−1). In addition, the maximum removal rate achieved by P. viridis was only 37% and occurred at M90 (Fig. 9b), one of its intermediate densities.
 |
Fig. 8 Concentration dynamics of NO3 (mg L−1) in the control and at various densities of G. verucosa (a) and P. viridis (b). |
 |
Fig. 9 The removal rates, averages and error bars of the four replications, of NO3 (%) in the control and at various densities of both G. verucosa (a) and P. viridis (b). |
3.5 Specific growth rate (SGR) of G. verucosa and P. viridis
The SGR of G. verucosa increased significantly (p < 0.01) with decreasing density (Tab. 1, Fig. 10i). The growth was slowest (1.6% day−1) at S200, while the higher SGRs at the three lower densities were not significantly different: 2.4% day−1 at S150, 2.55% day−1 at S100 and 2.6% day−1 at S50. Morphological observations showed that in the low densities, many young thallus appeared, whereas in the high densities, only a few young thallus emerged.
The SGR of green mussels was lowest (0.5% day−1) at M150, while it achieved 0.5% day−1 at M120, 0.6% day−1 at M90, and 0.8% day−1 at M60 (Fig. 10ii). The growth in the lowest density was significantly higher than that in the other three densities.
 |
Fig. 10 Specific Growth Rate (% day−1) of G. verucosa (a) and P. viridis (b) at four densities. Significant differences among the treatments are indicated by different lowercase letters. |
3.6 Water quality parameters
In all treatments, the water temperature ranged from 29 to 34 °C during the day and between 27 and 29 °C at night, and the salinity between 26 and 30 ppt (Tab. 2). However, the densities of both G. verucosa and P. viridis affected the pH, DO, transparency, and color of the water, although none of these trends were significant.
The lowest pH (7.5) was found in the treatments with the high densities: S150, S200, M120, and M150. The lowest DO content was obtained in the highest densities at night; during daytime the DO content remained in the same range. The water appeared muddier and more turbid in the tank with the lowest density of G. verucosa (transparency of 25 cm) and in the tank with the lowest density of P. viridis (transparency of 50 cm). The water in the tank with G. verucosa at S150 and in the tank with P. viridis at M90 was brownish-green and its transparency was around 60 cm. In the tank with S200 and the tank with M150, the water was so clear that the sediment bottom (at 0.8 m depth) was visible.
In water from M150, phytoplankton abundance was reduced significantly (p < 0.01). The phytoplankton abundance of the control and that of M60 and M90 was 8.2 × 106 ind. L−1, compared with 6.0 × 106 ind. L−1 for M120 and 5.2 × 106 ind. L−1 for M150 (Fig. 11).
The ranges of observed temperatures (T), dissolved oxygen (DO), pH, salinity, transparency and color of the water in the 8 treatments with 4 densities of both seaweed and green mussel.
 |
Fig. 11 The abundance of phytoplankton (×106) over time in the control and at various densities of P. viridis. |
4 Discussion
Renewing 10% of the pond water weekly mimicked real-life pond conditions and is necessary to avoid mortality. The water replaced the evaporation, and moreover carried the nutrients which are essential for maintenance and growth of macroalgae and mussel. All the water came from the same source and although the quality of the incoming water was not measured weekly, we assumed that this addition and its effects were the same for all tanks. Thus, experimental conditions remained the same for all treatments, but the reduction rates are thus relative and not absolute.
4.1 Total organic matter (TOM), total ammonia nitrogen (TAN), Nitrite (NO2), Nitrate (NO3)
In the best performing density, the capacity of P. viridis to remove TOM (consisting of dissolved, suspended, and colloidal OM) is greater than that of G. verucosa. P. viridis is an active filter feeder that ingests particles directly from the water column, whereas G. verucosa absorbs N and P from OM through the cell walls of the thallus.
Our maximum result, 8% TOM removal rate of S200, was lower than the 12% found by Soriano et al. (2009) for 20 g of Gracilaria birdiae in 10 L. Our findings on the impact of increasing stocking densities in our study align with those of other studies that have reported that removal rates vary significantly with stocking density and environmental conditions (Porrello et al., 2003; Bartoli et al., 2005; Cahill et al., 2010). Growing seaweed absorbs dissolved OM that has been degraded by micro-organisms, thus reducing OM levels in the water. In addition, seaweed also extracts colloidal particles from OM, thus making the waters clearer when seaweed densities are sufficiently high.
Our finding that TOM increased at high densities of green mussel agrees with that of Srisunot and Babel (2015). The increase is attributable to the mussels releasing soluble nutrient compounds (Tantanasarit et al., 2013), including the nutrients contained in their excreta. These causes of the increase in TOM at high densities had been confirmed in studies in mussel farms (Jones et al., 2001; Stadmark and Conley, 2011) and in an integrated culture of shrimp with blue mussel, Mytilus edulis: Van Khoi and Fotedar, 2012).
In our experiment, P. viridis removed up to 50%, TAN, up to 10% of NO2, and up to 40% of NO3. Srisunot and Babel (2016) found lower levels for TAN (23%) in an open water sea mussel farm. Our results for NO3, were higher than the 3.1% removal rate found by Tantanasarit and Babel (2014) but lower than the removal rates reported by Masilamani et al. (2001), which were 75% for NO3 and 73% for NO2. Previous studies support our findings that the capacity of green mussel to reduce the nutrient load by up to 380 mg yr−1 ind.−1, may vary due to size and environmental conditions (Irisarri et al., 2013; Srisunot and Babel, 2015). However, the trade-off is that each day the green mussels excrete close to 99 mg ind−1 of dry matter, and release about 2.5 mg NH4 +-N ind−1; these levels increase significantly under high densities (Srisunot and Babel, 2015). Under stressed conditions, such as high density, mussels increase their respiration rate as well as their NH4 +-N excretion (Christensen et al., 2003). Thus, at the two higher densities of P. viridis, M120 and M150, the higher total metabolic waste released to the water limited effective nutrient removal, and hence the levels of TOM, TAN, and NO2 increased.
G. verucosa's removal rate of TAN (67%) was higher than that reported by Carton-Kawagoshi et al. (2014) for Gracilaria sp. cultured in fish effluent (45%), but in that study and ours, the NO3 removal rate was similar (50%). The TAN removal rates we found for G. verucosa are in the same range as those found by Msuya and Neori (2002) and Nelson et al. (2001) for Gracilaria sp and that of Wei et al. (2017) for G. lemaneiformis, although their reported reduction efficiency was lower. However, at the end of our experiment, we found that TAN had increased in the G. verucosa treatment with the lowest density (S50); this density of G. verucosa might be too low to absorb the nitrogen in water added at regular intervals in our experiment, which was nutrient-rich. The highest removal rate of TAN (70%) was achieved by G. verucosa in S200, but at this high density, the NO2 level exceeded that in the control. This result demonstrates that there is an optimum density for the effective removal of both TAN and NO2. Nitrite increased at the highest density of G. verucosa, whereas NO3 decreased at all densities. At this density (S200), the DO level at night were at times as low as 2.6 ppm, thus reducing the activity of Nitrosomonas and Nitrobacter bacteria, a tandem needing oxygen to decompose ammonium through nitrite in nitrate in a two-step process (Watten and Sirbrell, 2006). Low levels of DO may be the cause of the increased nitrite concentration at high seaweed density, and as well play a role in the non-decrease of NO2 at the two highest densities of P. viridis (see Sect. 4.4).
4.2 Specific growth rates of G. verucosa and P. Viridis
In the three lower densities of seaweed, the SGR (2.3–2.6% day−1) was within the optimum range, that is, above 2% day−1 (Komarawidjaja and Kurniawan, 2008), but below the highest level (8.8%; range 1.8–8.8%) reported by Nelson et al. (2001). The seaweed's lower SGR at higher density (S200) is probably due to insufficient nutrient availability (Yang et al., 2006; Huo et al., 2012; Wei et al., 2017) and self-shading (Carton-Kawagoshi et al., 2014) which also inhibits the emergence of new thallus. Our results for temperature, salinity, and pH were within the optimal ranges of 20–30 °C, 17–40 ppt and 7–8.5, respectively (Komarawijaya and Kurniawan, 2008). Our findings confirm that in aquaculture, G. verucosa's SGR is mainly related to stocking density (Buschmann et al., 2001).
Mussels in higher densities compete for more available food and space for attachment (Srisunot and Babel, 2015), and indeed, we found that the green mussels grew less in higher densities (SGR at 0.5% day−1) than in lower densities (0.8% day−1). The latter rate is, however, lower than the rates reported by Srisunot and Babel (2015) and Tantanasarit et al. (2013), which were above 1% day−1in mussels cultured in open seawater. This discrepancy might be due to a difference in the relative nutrient concentration and abundance of plankton as food for mussels in our study.
The lower abundance of phytoplankton aligns with the lower SGR of green mussel under the two high densities. As argued earlier, at higher densities of mussel, less water is available per individual, which reduces food availability (Tantanasarit et al., 2013), and although the total filtration rate of food is high (Rajesh et al., 2001), the activity per individual decreases (Srisunot and Babel, 2015). This would account for the high SGR of P. viridis in lower densities.
4.3 Water quality parameters
Our finding that the highest nitrite concentration and the lowest pH (around 7.5) occurred in the treatments with the highest density of P. viridis align with that of (Boyd and McNevin, 2015), that the limited nitrification of the excreted ammonianitrogen and the decay by microorganism cause the pH to fall. The breakdown of OM increases oxygen consumption, which affects the nutrient cycle due to nitrogen depletion (Christensen et al., 2003; Vaquer-Sunyer and Duarte, 2008; Carlsson et al., 2010). During the night, the DO became at times low (2.6 and 2.9 mg/L−1) in the highest densities of Gracilaria sp and P. viridis (S200 and M120 resp.), which − as noted above −hampered nitrification in the highest densities of both. For the lower densities of G. verucosa and P. viridis, the levels of TAN and nitrite remained favorable for cultivation.
Given the results of this study, our next step would be to improve water quality for shrimp culture by combining the medium densities of G. verucosa and the lower densities of P. viridis. In addition to our aim of establishing the optimal removal rates of organic waste, we would like to ascertain the optimal growth of these by-products, that is, seaweed and green mussels, so that we could diversify the farmers' sources of revenue.
5 Conclusion
Our study shows that P. viridis is more effective than G. verucosa in reducing TOM: the RRs were 40% and 10%, respectively. However, G. verucosa is more effective than P. viridis in removing TAN: RRs of about 60% and 50% for G. verucosa versus RRs of 50% and 30% for P. viridis. Low densities of G. verucosa (50 and 100 g m−2) and P. viridis (60 and 90 g m−2) resulted in the highest RRs of NO2, 50% and 10%, respectively. In contrast, the highest densities of both increased NO2 instead of removing it. The growth rates of G. verucosa were significantly higher at the three lower densities (50, 100, and 150 g m−2) than at 200 g m−2. The density effect was not significant for P. viridis, but its growth was lowest at the highest density (150 g m−2). In our future research on integrated multi-trophic shrimp aquaculture, to optimize the removal rates of OM and nitrogen in tanks with shrimp, we intend to combine medium levels of G. verucosa (around 100 g m−2) and lower levels of P. viridis (around 60 g m−2).
Conflict of interest
The authors declare that they have no conflict of interest. All grants came from non-commercial partners.
Ethical approval
The research reported in this article did not involve any studies performed by the authors on animals, given that most advanced ethical regulations exclude crustaceans and shellfish.
Acknowledgments
We gratefully acknowledge Dr. Marc Verdegem of the Aquaculture and Fisheries group, Wageningen University for reviewing the statistical analysis of this study, Dr. Lorna M. Calumpang for editing this paper and Dr. Joy Burrough-Boenisch for reviewing the scientific English. We also extend our deep gratitude to those who funded this study: the Non-APBN Research from the Faculty of Fisheries and Marine Science, Diponegoro University, the NWO-WOTRO in the framework of the Project to design Aquaculture for Supporting Mangrove restoration in Indonesia (PASMI), the Eco-shape Foundation based in the Netherlands and the NGO Wetlands International Indonesia, and the Program Enhancing International Publication from the Indonesian Ministry of Research, Technology and High Education.
References
- Aliah R. 2012. Modelling of integrated multi trophic aquaculture on the north coast of Karawang, west Java. J Environ Eng 13: 47–52. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Waste Water, 22nd edn, American Public Health Association, Washington DC, 2012, p. 1496. [Google Scholar]
- Bartoli M, Nizzoli D, Naldi M, Vezzulli L, Porrello S, Lenzi M, Viaroli P. 2005. Inorganic nitrogen control in wastewater treatment ponds from a fish farm (Orbetello, Italy): denitrification versus Ulva uptake. Mar Pollut Bull 50: 1386–1397 . [Google Scholar]
- Boyd CE, McNevin AA. 2015. Aquaculture, Resource Use and the Environment, Wiley Blackwell, Canada. [Google Scholar]
- Busacker GP, Adelman TR, Goolish EM. Growth, in: C.B. Schreck, P.B. Moyle (Eds.), Method for Fish Biology, American Fisheries Society, Bathesda, Maryl, 1990, pp. 363–377 . [Google Scholar]
- Buschmann A, Troell M, Kautsky N. 2001. Integrated algal farming: a review. Cah Biol Mar 42: 83–90 . [Google Scholar]
- Cahill PL, Hurd LC, Lokman M. 2010. Keeping the water clean − Seaweed biofiltration outperforms traditional bacterial biofilms in recirculating aquaculture. Aquaculture 306: 153–159 . [Google Scholar]
- Carlsson MS, Glud RN, Petersen JK. 2010. Degradation of mussel (Mytius edulis) fecal pellet released from hanging long-line upon sinking and after settling at the sediment. Can J Fish Aquat Sci 67: 1376–1387 . [Google Scholar]
- Carton-Kawagoshi RJ, Elle BJ, Corre Jr. V, Satoh S, Notoya M, Fujita D. 2014. Low water exchange Gracilariopsis bailiniae, Zhang & Xia, culture in intensive milkfish culture effluents: effects of seaweed density on seaweed production and effluent treatment. Aquacult Int 22: 573–584 . [Google Scholar]
- Chaiyakum K, Tanwilai D. Experiment on using of Green Mussel, Mytilus sp. and Seaweed, Gracilariafisheri for Biological Wastewater Treatment from Intensive Culture of Tiger Shrimp Ponds, National Institute of Coastal Aquaculture, Songkhla, Thailand. Technical paper 6, 1992. [Google Scholar]
- Christensen PB, Glud RN, Dalsgaard T, Gillispie P. 2003. Impacts of longline mussel farming on oxygen and nitrogen dynamics and biological communities of coastal sediments. Aquaculture 218: 567–588 . [Google Scholar]
- Filgueira R, Strople LC, Sttohmeimer T, Rastrick S, Strand O. 2019. Mussels or tunicates: that is the question. Evaluating efficient and sustainable resource use by low-trophic species in aquaculture settings. J Clean Prod 231: 132–143. [Google Scholar]
- Golez NV, Mallare MT, Guanzon N, Hurtado AQ, Jaspe J. A study on the integrated biological processes for treating intensive shrimp effluents. Sediment Quality Assessment Program, Chicago, IL, 16–18 October 2002. [Google Scholar]
- Haamer J. 1996. Improving water quality in a eutrophied fjord system with mussel farming. Ambio 25: 356–362. [Google Scholar]
- Huo YZ, Wu HL, Chai ZY, Xu SN, Han F, Li D, He PM. 2012. Bioremediation efficiency of Gracilaria verrucosa for an integrated multi-trophic aquaculture system with Pseudosciaena crocea in Xiangshan harbor, China. Aquaculture 326–329: 99–105 . [Google Scholar]
- Irisarri J. Reiriz MJF, Robinson SMC, Cranford PJ, Labarta U. 2013. Absorption efficiency of mussels Mytilus edulis and Mytilus galloprovincialis cultured under Integrated Multi-Trophic Aquaculture conditions in the Bay of Fundy (Canada) and Ría Ares-Betanzos (Spain). Aquaculture 381–391: 182–192 . [Google Scholar]
- Jones AB, Dennison WC, Pretson NP. 2001. Integrated treatment of shrimp effluent by sedimentation, oyster filtration and macroalgal absorption: a laboratory scale study. Aquaculture 193: 155–178 . [Google Scholar]
- Komarawidjaja W, Kurniawan DA. 2008. Filtration rate of seaweed (Gracilariasp) to the concentration of orthophosphate (P2O5). J Environ Eng 9: 180–183. [Google Scholar]
- Kutty MN. 1987. Site Selection for Aquaculture, Chemical features of water. African Regional Aquaculture Centre, Port Harcourt, Nigeria. [Google Scholar]
- Marinho-Soriano ESO, Nunes MAA, Carneiro, Pereira DC. 2009. Nutrients' removal from aquaculture wastewater using the Macroalgae Gracilaria birdiae . Biomass Bioenerg 33: 327–331. [Google Scholar]
- Masilamani JG, Azariah J, Nandakumar K, Jesudoss KS et al. 2001. Excretory products of green mussel (Perna viridis L.) and their implications on power plant operation. Turk J Zool 25: 117–125 . [Google Scholar]
- Ministry of Marine Affairs and Fisheries (MMAF) of Republic Indonesia. 2013. Government Regulation No. 75. about Rearing tiger shrimp (Penaeus monodon) and vannamei shrimp (Litopenaeus vannamei). [Google Scholar]
- Msuya FE, Neori A. 2002. Ulva reticulata and Gracilariacrassa: macroalgae that can biofilter effluent from tidal fishpons in Tanzania. Western Indian Ocean. J Mar Sci 1: 117–126 . [Google Scholar]
- National Standarization Agency of Indonesia. 2004. SNI: 06-6989.22. Water and water waste: Chapter 22. Organic matter test using spectrophotometry by titration of permanganate. [Google Scholar]
- National Standarization Agency of Indonesia. 2011. SNI: 6989.79. Water and water waste: Chapter 31. Nitrate test using spectrophotometry by cadmium reduction method. [Google Scholar]
- Nelson S, Glenn E, Moore D, Walsh T, Fitzsimmons K. 2001. Use of an edible red seaweed to improve effluent from shrimp farms. Environmental Research Laboratory, Univ. Arizona. Tucson. AZ. [Google Scholar]
- Neori A, Chopin T, Troel M, Buschmann AH et al. 2004. Integrated aquaculture: rationale, evolution and state of the art emphasizing seaweed biofiltration in modern mariculture. Aquaculture 231: 361–391 . [Google Scholar]
- Pandjara B, Hendrajat EA, Surwoyo HS. 2010. Utilization of biofilter in tiger shrimp culture in abandon pond. Prosiding Forum Inovation. Aquac Technol 221–131 . [Google Scholar]
- Pena-Rodríguez A, Magallón-Barajas FJ, Cruz-Suárez LE, Elizondo González R, Moll B. 2017. Effects of stocking density on the performance of brown shrimp Farfantepenaeuscaliforniensis cocultured with the green seaweed Ulva clathrata. Aquac Res 48: 2803–2811 . [Google Scholar]
- Porrello S, Mauro L, Persia E, Tomassetti P, Finoia MG. 2003. Reduction of aquaculture wastewater eutrophication by phytotreatment pond systems: dissolved and particulate nitrogen and phosphorous. Aquaculture 219: 515–529 . [Google Scholar]
- Rabiei R, Phang S, Yeong H, Lim P, Ajdari D, Zarshenas G, Sohrabipour J, 2014. Bioremediation efficiency and biochemical composition of Ulva reticulata Forsskål (Chlorophyta) cultivated in shrimp (Penaeus monodon) hatchery effluent. Iran J Fish Sci 13: 621–639. [Google Scholar]
- Rahmaningsih S. 2012. Technology using seaweed as a natural biofilter in brackish-water to reduce the disease attack in Litopenaeus vannamei . Fish Technol Oceanogr 2: 11–16 . [Google Scholar]
- Rajesh KV, Mohamed KS, Krippa V. 2001. Influence of algal cell concentration, salinity and body size on the filtration and ingestion rate of cultivable India bivalves. Indian J Mar Sci 30: 87–92 . [Google Scholar]
- Statistic of Central Java. 2020. https://jateng.bps.go.id/statictable/2020/07/22/1953/produksi-dan-nilai-produksi-perikanan-budidaya-menurut-kabupaten-kota-dan-komoditas-utama-di-provinsi-jawa-tengah-2018x.html [Google Scholar]
- SNI (Standard National Indonesia), 2004. Water and Wastewater − Methodology for calculating permanganate titration. National Standarization Department. Indonesia. [Google Scholar]
- Susilowati T, Hutabarat J, Anggoro S, Zainuri M. 2014. The improvement of the survival, growth of naname shrimp (Litopenaeus vannamei) and seaweed (Gracilaria verucosa) based on polyculture cultivation. IJMARCC 1: 6–11 . [Google Scholar]
- Srisunot C, Babel S. 2016. Estimating the carrying capacity of green mussel cultivation by using net nutrient removal model. Mar Pollut Bull 112: 235–243 . [Google Scholar]
- Srisunot C, Babel S. 2015. Uptake, release, and absorption of nutrients into the marine environment by the green mussel (Perna viridis). Mar Pollut Bull 97: 285–293 . [Google Scholar]
- Stadmark J, Conley DJ. 2011. Mussel farming as a nutrient reduction measure in the Baltic Sea: consideration of nutrient biogeochemical cycles. Mar Pollut Bull 62: 1385–1388. [Google Scholar]
- Tantanasarit C, Babel S. 2014. Simulation of net nutrients removal by green mussel (Perna viridis) in Estuarine and Coastal Areas. Int J Environ Ecol Eng 8: 121–113 . [Google Scholar]
- Tantanasarit C, Babel S, Englande AJ, Meksumpun S. 2013. Influence of size and density on filtration rate modelling and nutrient uptake by green mussel (Perna viridis). Mar Pollut Bull 68: 38–45 . [Google Scholar]
- Tendencia EA. 2007. Poluculture of green mussel, brown mussles and oysters with shrimp control luminous bacterial disease in a simulated culture system. Aquaculture 272: 188–191. [Google Scholar]
- Tendencia EA, Fermin AC, dela Pena MR, Choresca Jr CH. 2006. Effect of Epinephelus coioides, Chanoschanos, and GIFT tilapia in polyculture with Penaeus monodon on the growth of the luminous bacteria Vibrio harveyi . Aquaculture 253: 48–56. [Google Scholar]
- Van Khoi L, Fotedar R. 2012. Integration of blue mussel (Mytilus edulis Linnaeus, 1758) with western king prawn (Penaeus latisulcatus Kishinouye, 1896) in a closed recirculating aquaculture system under laboratory conditions. Aquaculture 345–355: 84–90. [Google Scholar]
- Vaquer-Sunyer R, Duarte CM. 2008. Treshold of hypoxia for marine biodiversity. Proc Natl Acad Sci U S A 105: 15452–15457. [PubMed] [Google Scholar]
- Watten BJ, Sirbell PL. 2006. Comparative performance of fixed film biological filters: application of reactor theory. Aquac Eng 34: 193–213 . [Google Scholar]
- Wei Z, You J, Wu H, Yang F, Long L, Liu Q. 2017. Bioremediation using Glacilaria lemaneiformis to manage the nitrogen and phosphorous balance in an integrated multi-trophic aquaculture system in Yantian Bay, China. Mar Pollut Bull 121: 313–319. [Google Scholar]
- Yang YF, Fei XG, Song JM, Hu HY, Wang GC, Chung IK. 2006. Growth of Gracilaria lemaneiformis under different cultivation conditions and its effects on nutrient removal in Chinese coastal waters. Aquaculture 254: 248–255 . [Google Scholar]
- Yuniarsih E, Nirmala K, Radiarta IN. 2014. The removal rate of nitrogen and phosphor at seaweed culture with Integrated Multi Trophic Aquaculture in Gerupuk Bay, Lombok, Nusa Tenggara Barat. J Riset Aquac 9: 487–500 . [Google Scholar]
Cite this article as: Widowati LL, Budi Prayitno S, Rejeki S, Elfitasari T, Purnomo PW, Ariyati RW, Bosma RH. 2021. Organic matter reduction using four densities of seaweed (Gracilaria verucosa) and green mussel (Perna viridis) to improve water quality for aquaculture in Java, Indonesia. Aquat. Living Resour. 34: 5
All Tables
Results of the two-way ANOVA testing the influence of days, density, and the interaction on TOM, TAN, NO2, NO3, and SGR of G. verucosa and P. viridis.
The ranges of observed temperatures (T), dissolved oxygen (DO), pH, salinity, transparency and color of the water in the 8 treatments with 4 densities of both seaweed and green mussel.
All Figures
|
Fig. 1 G. verucosa (a) and P. viridis (b) were cultured by the hanging method. |
| In the text | |
|
Fig. 2 Concentration dynamics of TOM (mg L−1) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
| In the text | |
|
Fig. 3 The removal rates, averages and error bars of the four replications, of TOM (%) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
| In the text | |
|
Fig. 4 Concentration dynamics of TAN (mg L−1) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
| In the text | |
|
Fig. 5 The removal rates, averages and error bars of the four replications, of TAN (%) in the control and at four densities of both G. verucosa (a) and P. viridis (b). |
| In the text | |
|
Fig. 6 Concentration dynamics of NO2 (mg L−1) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
| In the text | |
|
Fig. 7 The removal rates, averages and error bars of the four replications, of NO2 (%) in the control and at four densities of G. verucosa (a) and P. viridis (b). |
| In the text | |
|
Fig. 8 Concentration dynamics of NO3 (mg L−1) in the control and at various densities of G. verucosa (a) and P. viridis (b). |
| In the text | |
|
Fig. 9 The removal rates, averages and error bars of the four replications, of NO3 (%) in the control and at various densities of both G. verucosa (a) and P. viridis (b). |
| In the text | |
|
Fig. 10 Specific Growth Rate (% day−1) of G. verucosa (a) and P. viridis (b) at four densities. Significant differences among the treatments are indicated by different lowercase letters. |
| In the text | |
|
Fig. 11 The abundance of phytoplankton (×106) over time in the control and at various densities of P. viridis. |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.