| Issue |
Aquat. Living Resour.
Volume 34, 2021
|
|
|---|---|---|
| Article Number | 10 | |
| Number of page(s) | 8 | |
| DOI | https://doi.org/10.1051/alr/2021009 | |
| Published online | 29 April 2021 | |
Research Article
Synbiotic containing Kappaphycus alvarezii, Spirulina sp. and Halomonas alkaliphila improves survival, growth and vibriosis resistance in whiteleg shrimp (Litopenaeus vannamei) post-larval culture
1
Microbial Biotechnology Research Group, School of Life Sciences and Technology, Institut Teknologi Bandung, Jalan Ganesa No. 10 Bandung, Indonesia
2
Department of Microbiology, School of Life Sciences and Technology, Institut Teknologi Bandung, Jalan Ganesa No. 10 Bandung, Indonesia
* Corresponding authors: This email address is being protected from spambots. You need JavaScript enabled to view it.
Handling Editor: Domenico Caruso
Received:
29
May
2020
Accepted:
11
March
2021
Abstract
The high losses in shrimp production due to mortality caused by Vibrio opportunistic pathogens still constitute a significant challenge in the shrimp industry. Synbiotic feed supplementation appears to be a promising control strategy to maintain healthy shrimp stock. In this study, the effects of synbiotic-containing prebiotic seaweeds Kappaphycus alvarezii and Spirulina sp. as well as probiotic Halomonas alkaliphila were evaluated on the survival, growth, and vibriosis of Litopenaeus vannamei during the post-larval stage. Five different feeds were tested: commercial feed, prebiotics K. alvarezii and Spirulina sp.-supplemented feed, and synbiotic-supplemented feed using K. alvarezii, Spirulina sp. and probiotic H. alkaliphila with different concentrations of 108, 109, and 1010 CFU.kg−1. Following 14 days after the feeding test, the highest shrimp survival (91.46 ± 0.05%) was obtained in the treatment group fed with synbiotic-supplemented feed containing 0.375% K. alvarezii, 0.125% Spirulina sp., and H. alkaliphila at 109 CFU.kg−1 (p < 0.05). A 7-day challenge test against opportunistic bacteria Vibrio harveyii was then performed using three treatment groups: (1) synbiotic, containing 0.375% K. alvarezii, 0.125% Spirulina sp., and H. alkaliphila at 109 CFU.kg−1; (2) prebiotic, with 0.375% K. alvarezii, 0.125% Spirulina sp.; and (3) control, using commercial feed. The highest shrimp survival of (79.9 ± 0.05%) was found in the synbiotic treatment group, followed by the prebiotic and control treatment groups (p < 0.05). Overall results suggested that synbiotic-supplemented feed containing 0.375% K. alvarezii, 0.125% Spirulina sp., and H. alkaliphila at 109 CFU.kg−1 significantly improved shrimp survival even when challenged with V. harveyii. Thus, this synbiotic can be potentially applied as an alternative biocontrol strategy against vibriosis in intensive shrimp post-larval culture.
Key words: Halomonas / Kappaphycus / Shrimp / Spirulina / Synbiotic / Vibrio
© EDP Sciences 2021
1 Introduction
With an average growth of 8.6% per year, aquaculture shows promising results in meeting the global demand in fish products, which is expected to increase about 50% by 2050 (Boyd and Nevin, 2015; FAO, 2018). The whiteleg shrimp Litopenaeus vannamei is one of the main species used in aquaculture, particularly in South-East Asia, which makes up 65% of the global shrimp production (FAO, 2019). However, high losses in shrimp production due to mortality caused by viruses and pathogenic bacteria are still a significant challenge to meeting global production quotas (Chiu et al., 2007). One of the common diseases found in shrimp is vibriosis, a disease caused by bacteria of the Vibrio genus, such as Vibrio harveyii. This disease is known to cause 70–90% of shrimp mortality during the post-larval stage, when its concentration in shrimp culture water reaches up to 106 CFU.ml−1 (FAO, 2014; Suantika et al., 2017; Suantika 2018b). The total Vibrio sp. population in the shrimp culture water in optimal conditions has been reported to be 103–105 CFU.ml−1 (Suantika et al., 2017, 2018a,b,c). However, opportunistic Vibrio may become pathogenic in specific environmental conditions, e.g. shrimp overcrowding stress in super-intensive aquaculture (Brock and Lightner, 1990).
The emergence of antibiotic-resistant strains of pathogenic bacteria has become a central issue in aquaculture, so alternatives for disease prevention and control need to be developed to address this emerging resistance (Smith, 2008). Formulating shrimp feed supplements to increase shrimp survival rate, growth, and disease resistance has − so far − been the most popular approach (Situmorang et al., 2020). In the last decade, the development of synbiotics, a mixture of pre- and probiotics, has been intensively studied for their potential application as feed supplements in L. vannamei culture (Oktaviana et al., 2014; Arisa et al., 2015; Huynh et al., 2018). Probiotics are live microbes which impart beneficial effects on host cells (Das et al., 2017). In L. vannamei culture, these benefits include the enhanced production of digestive enzymes in the shrimp gut, controlling the dominance of pathogenic bacteria, activating the intestinal immune system, and improving nutrient absorption (Zhou et al., 2009; Ramezani et al., 2014). Prebiotics are non-digestible food compounds that can selectively stimulate the growth or activity of beneficial bacteria in the digestive tract (Arisa et al., 2015). Several studies have shown that L. vannamei fed with prebiotic supplements show increased antimicrobial activity, enhanced phagocytic capacity, and less attachment of pathogenic bacteria in the gut (Anuta et al., 2014; Hardjani et al., 2017). Synbiotics alludes to synergism, and this term should be reserved for products in which the prebiotic selectively favor the probiotic organism(s) (Cencic and Chingwaru, 2010). Synbiotics were developed to overcome the possible survival difficulties for probiotics. A more efficient stimulating effect of the growth of probiotics and ubiquitous bacteria contribute on maintaining the intestinal homeostasis and a healthy body (Pandey et al., 2015).
Recently, the red seaweed Kappaphycus alvarezii has been studied as a potential prebiotic in the production of shrimp feed supplements (FAO, 2014; Suantika et al., 2017; Suantika 2018b). K. alvarezii is one of the main aquatic plants produced in aquaculture, with a global total production of 1.6 million tons in 2016 (FAO, 2018). K. alvarezii contains kappa-carrageenan, a potential immunostimulant which may improve shrimp immune response against pathogens (Hardjani et al., 2017; Suantika et al., 2017). However, this species lacks some amino-acid and essential fatty acid content and thus needs to be supplemented with other substances prior to its use as a shrimp feed supplement. The cyanobacteria Spirulina sp. can be used to complement K. alvarezii, as it contains 60–70% protein and the essential amino acids present are around 47% of total protein weight that are important in shrimp diets (Glencross et al., 2020; Ibrahem et al., 2013). The following amino acids required by shrimp for the optimal growth were provided in Spirulina: isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophane, and valine (Glencross et al., 2020).
Several probiotic bacteria have been investigated in L. vannamei diet supplementation, growth, and disease control (Zhang et al., 2009; Chandran et al., 2014). In shrimp culture, Halomonas sp. is used as a probiotic and has been shown to increase shrimp resistance to the white spot syndrome virus (Zhang et al., 2009). In general, the probiotic products should have a minimum concentration of 106 CFU.ml−1 or gram and a total of around 108–109 probiotic microorganisms should be consumed daily for the probiotic effect to be transferred to the consumer (Kechagia et al., 2013). Our study aimed to investigate the effect of a dietary synbiotic containing K. alvarezii and Spirulina sp. prebiotics and local probiotic isolates of H. alkaliphila with optimal concentration for the survival, growth, and vibrosis resistance of post-larval shrimp culture.
2 Materials and methods
2.1 Probiotic production
H. alkaliphila, isolated from shrimp, was grown on Luria Bertani medium (Oxoid, Indonesia) (30, 25 °C) for 12 h. Bacterial cells were harvested through centrifugation (445 × g, 5 min, 25 °C); bacterial pellets were then rinsed using sterile distilled water to remove the remaining medium and freeze-dried (FD) using Freeze Dryer VD-800F (Taitec, Japan) for 24 h. The concentration of viable bacteria from the resultant powder after freeze-drying (CFU.gram FD−1) was measured using the total plate count (TPC) method with a spread technique on a nutrient agar medium (Oxoid, Indonesia) with salinity 30 (Cappuccino and Sherman, 2014). The amount (g of FD) of probiotic powder addition on feed (P) was calculated by: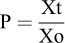 where Xt was the final probiotic concentration in the feed (CFU) and Xo was the concentration of the bacterial powder (CFU.g FD−1).
where Xt was the final probiotic concentration in the feed (CFU) and Xo was the concentration of the bacterial powder (CFU.g FD−1).
2.2 Synbiotic feed production and formulation
Synbiotic feed was produced through the addition of synbiotic products into commercial (basal) feed. The commercial feed (Feng Li®, Indonesia) contained 41% protein, 7% lipid, 3% fiber, 13% ash, and 10% water. In this study, synbiotic feeds were produced by adding the prebiotic K. alvarezii (Helos®, PT. Gapura Akua Kultiva, Indonesia) at a concentration of 0.375% and Spirulina sp. (CERES®, Indonesia) at a concentration of 0.125% to the commercial feed, as well as H. alkaliphila powder at three different concentrations: 108 CFU.kg−1 feed (S8 group), 109 CFU.kg−1 feed (S9 group); and 1010 CFU.kg−1 feed (S10 group). The prebiotic feed (P group) was formulated by adding the prebiotic K. alvarezii and Spirulina sp. in the concentrations stated above, without H. alkaliphila. The commercial feed control (C group) was fed with 100% commercial feed without added synbiotics. Feed was stored in a dry place at room temperature (25 °C).
2.3 Proximate analyses
Analyses were performed at Saraswanti Indo Genetech (Bogor, Indonesia) to determine the total energy per biomass and total energy from fat, as well as water, ash, total lipid, protein, and total carbohydrate content in all five diets. Ash content was measured by heating a sample to 600 °C until a constant weight was obtained (Situmorang et al., 2020). Protein content was measured using the Kjeldahl method (Maehre et al., 2018), and the lipid content was measured using the Weibull method (Situmorang et al., 2020). Carbohydrate content (including nitrogen-free extract and crude fiber) was calculated by subtracting the other elements from the total weight (FAO, 2014).
2.4 Experimental set-up and feeding test
A batch culture system using 15 10-L tanks equipped with aeration lines and water heaters and using seawater from Indramayu, Jawa Barat, Indonesia to culture shrimps. The seawater was sterilized using sodium hypochlorite (Bayclin®, Indonesia), then neutralized using sodium thiosulfate. Initial culture system conditions were as follows: salinity, 30–33; temperature, 30–32 °C; dissolved oxygen, 4–7 mg.L−1 (Suantika et al., 2013). A total of 9750 specific pathogen-free (SPF) 5-day-old shrimp in the post-larval stage (PL5) with an average initial mean (± SD) body weight of 2.74 ± 0.23 mg and an average initial body length (±SD) of 0.64 ± 0.11 cm (random sampling from 20 shrimps) were stocked in 15 culture tanks at a high density of 650 PL per tank. All treatments were tested in triplicates over 14 days. Shrimp were fed four times a day 10% of their initial body weight and adjustments were made every day, assuming the shrimps gained 10% weight daily (Suantika et al., 2018a,b,c). During the experimental period, a 10% water renewal and debris/sediment siphoning were performed three times a week to maintain optimal water quality parameters. Water physicochemical parameters, including pH, temperature, dissolved oxygen (DO), and salinity level were measured every two days. NH4+ and NO2− levels were also measured every two days using Nessler, diazotation, and nitrate HCl methods (Suantika et al., 2018a,b,c). The synbiotic treatment enabling the best shrimp survival was selected to undergo a bacterial challenge test using opportunistic pathogen V. harveyii.
2.5 Vibrio harveyii production and challenge test
After 14 days of feeding test, a bacterial challenge test for the C, P, and S9 treatment groups was performed for 7 days using V. harveyi obtained from Balai Besar Perikanan Budidaya Air Payau Jepara, Central Java, Indonesia. V. harveyii was cultured in Luria Bertani Broth medium (Oxoid, Indonesia) (salinity 30, temperature 25 °C) for 12 h. After incubation, the density of the bacterial culture was determined by measuring turbidity with a spectrophotometer (Genesys 20, Termospectronic) at a wavelength of 550 nm. V. harveyii culture (10 ml) at a concentration of 1010 CFU.ml−1 was harvested by centrifuging at 89 × g for 10 min and washed twice in its culture medium, then washed once using shrimp culture water before being added to each respective tank. The final concentration of V. harveyii in the tank was 106 CFU.ml−1. For each experimental feed in the challenge test, a non-exposed group without Vibrio (non-challenged) was tested and compared to the groups infected with Vibrio. Each treatment was performed in triplicates over seven days.
2.6 Bacterial enumeration of shrimp and culture water samples
Three post-larval shrimps were collected from each tank at day 0, 7, and 14 during the feeding test, euthanized in an ice-cold 5 g L−1 benzocaine solution (Sigma, E-1501), then rinsed with sterile seawater. Shrimp samples from the same tank were pooled and homogenated using a micro pestle with sterile NaCl 0.85% in a 1.5-ml microtube. Additionally, 50 ml of culture water was also collected from each tank. Shrimp homogenate was diluted 100× using NaCl 0.85%; the culture water sample was similarly diluted. The number of heterotrophic bacteria in shrimp and water sample were recorded using the total plate count (TPC) method with spread technique using 100 μL of diluted shrimp gut homogenate on natrium agar medium (Oxoid, Indonesia) with salinity 30, respectively. Inoculated agar plates were incubated at room temperature (25 °C) for 24 h before the number of grown colonies was counted (Cappuccino and Sherman, 2014).
The initial and final concentration of Vibrio sp. in water and shrimp cultures from the challenged groups also was measured using the TPC method with a spread technique on thiosulfate-citrate-bile salts-sucrose (TCBS) agar medium (Oxoid, Indonesia) with salinity 30 (Cappuccino and Sherman, 2014).
The Shannon-Weaver index to measure bacterial diversity in shrimp and culture water samples was calculated using the following formula (Lemos et al., 2011):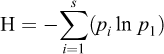
where s is the number of operational taxonomic unit (OTUs) and p is the proportion of the community represented by OTU i. The Shannon-Weaver index estimates species richness species evenness within a given sample (Lemos et al., 2011).
2.7 Bacterial identification
The dominant bacterial culture observed from shrimp culture during feeding test was purified and identified by bacterial DNA extraction, polymerase chain reaction amplification (27F/1492R primer), and bacterial rRNA 16S gene sequencing (785F/907R primer) at Macrogen, Korea. The sequence homology search for test sequences was performed using Nucleotide Blast versus GenBank and Ribosomal Database Project data. Results were validated via a phylogenetic analysis using the Winclada program and the Ratchet method (Island Hopper) (Suantika et al., 2017).
2.8 Growth and survival measurements
The average survival percentage, weight gain, and length increase following the 14-day feeding experiment and 7-day challenge test of each treatment groups were calculated as follows:
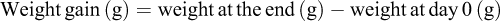
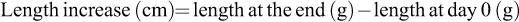
2.9 Statistical analysis
Survival data were first normalized using an arcsine transformation before statistical analysis (Suantika et al., 2018a,b,c). To evaluate the differences between treatment groups, data on growth and survival parameters were analyzed using one-way ANOVA and followed by Duncan posthoc Test with 95% confidence intervals. All statistical analyses were performed using SPSS® Version 24.0.
3 Results
3.1 Probiotic concentration and effect of its addition into the experimental feed
The probiotic powder was produced through the freeze-drying method, a commonly used technique for the production of dried probiotics. Cells are first frozen to below the critical temperature of the formulation, then dried by sublimation under high vacuum in two phases: primary drying, during which unbound water is removed; and secondary drying, during which the bound water is removed (Savani et al., 2010; Jalali et al., 2012). The TPC in the freeze-dried bacterial powder showed that viable H. alkaliphila was found at a concentration of 2.0 × 1010 CFU.g FD−1. Three different synbiotic feeds were made by adding 5.19, 51.9, and 518.7 mg of freeze-dried H. alkaliphila into commercial feed for S8, S9 and S10, respectively.
3.2 Proximate dietary analysis
The percentages of each component from the three different diets used in this study are presented in Table 1. Generally, the synbiotic-supplemented feed contained a higher amount of carbohydrate compared to commercial feed (i.e., the control), while the S9 diet had the highest total energy (360.68 kcal.100 g−1).
Proximate analysis results of commercial feed (C), prebiotic-supplemented feed (P), synbiotic-supplemented feed using Halomonas alkaliphila 108 CFU.kg−1 (S8), Halomonas. alkaliphila 109 CFU.kg−1 (S9), and Halomonas alkaliphila 109 CFU.kg−1 (S10) used in this experiment, with the optimum range values provided for comparison.
3.3 Water quality parameters
All water quality parameters were in the optimum range for shrimp cultivation (Suantika et al., 2018a,b,c), including dissolved oxygen (4.0–6.0 mg.L−1), pH (7.0–8.5), temperature (29–32 °C), salinity (30–35), ammonium (0.01–0.15 mg.L−1), and nitrite (0.1–1.1).
3.4 Analysis of shrimp biological parameters in feeding test
Following the 14 days culture period, the survival of L. vannamei in the synbiotic (S9) and prebiotic (P) group was significantly higher than the control group (p < 0.05). The survival S8 and S10 (71.7% and 66.2%, respectively) were not significantly different from C (65.7%) or P (78%; p > 0.05; Tab. 2). The highest survival rate was 91.5% in the S9 group, which was significantly higher than the control (survival rate: 65.7%; p < 0.05). No significant differences in the changes in average body weight (+3–6 mg) and length (+0.23–0.34 cm) were detected between the treatment groups (Tab. 2). The S9 and P groups were subsequently be used in the challenge test against V. harveyii and compared to the C treatment group as a control.
The effect of the 14-day feeding test on Litopenaeus. vannamei using control commercial feed (C); prebiotic-supplemented feed (P); and synbiotic-supplemented feed with Halomonas alkaliphila 108 CFU.kg−1 (S8), 109 CFU.kg−1 (S9), and 1010 CFU.kg−1 (S10).
3.5 Shrimp biological parameters in the challenge test against V. harveyii
In the challenge test, the highest survival of L. vannamei was obtained by the S9 group in both non-challenged and challenged conditions (p < 0.05). The survival of L. vannamei in the P and C groups significantly decreased with the addition of a lethal dose of V. harveyii (i.e., 106 CFU.ml−1; p < 0.05). However, there was no reduction of shrimp survival in the S9 group or synbiotic treatment even in the post-challenge test. In general, no significant differences in changes in average body weight (+0.02–0.05 g) and length (+0.15-0.25 cm) were found between treatments groups (Tab. 3).
Results of the 7-day challenge test against Vibrio harveyi on post-larval Litopenaeus vannamei with control commercial feed (C), prebiotic-supplemented feed (P), and synbiotic-supplemented feed with Halomonas alkaliphila 109 CFU.kg−1 (S9).
3.6 Analysis of dominant bacterial colonies during the feeding test
The Shannon-Weaver index in the synbiotic treatment was higher than that of the control, suggesting its bacterial community was diverse and abundant (Fig. 1). Pre-dominant bacterial isolates showed differences between sampling time points and between treatments. The synbiotic treatment resulted in more variation in the diversity of dominant bacterial isolates in both the water and shrimp samples, including species like Bacillus cereus, Pseudoasteromonas piscicida, and Meridianimaribacter flavus (Fig. 2).
 |
Fig. 1 The Shannon-Weaver index of the bacterial community in shrimp culture and water culture water during the feeding test at day 0 (T0), day 7 (T1), and day 14 (T2) during the feeding test using commercial feed (C), prebiotic-supplemented feed (P), synbiotic-supplemented feed using Halomonas alkaliphila 108 CFU.kg−1 (S8), Halomonas alkaliphila 109 CFU.kg−1 (S9), and Halomonas alkaliphila 109 CFU.kg−1 (S10). |
 |
Fig. 2 Percentage of dominant bacterial community in shrimp culture (a) and water culture (b) at day 0 (T0), day 7 (T1), and day 14 (T2) during the feeding test using commercial feed (C), prebiotic-supplemented feed (P), synbiotic-supplemented feed using Halomonas alkaliphila 108 CFU.kg−1 (S8), Halomonas alkaliphila 109 CFU.kg−1 (S9), and Halomonas alkaliphila 109 CFU.kg−1 (S10). |
3.7 Concentration of Vibrio sp. in the challenge test
In general, Vibrio sp. in cultivation systems reaches the lethal dose for shrimps at ∼106 CFU.ml−1. The prebiotic and synbiotic treatments can reduce the total concentration of Vibrio sp. in both the shrimp and water samples; however, the synbiotic treatment was the most effective in this regard: achieving a reduction from 106 CFU.ml−1–102 CFU.ml−1 in culture water (Tab. 4).
The total number of Vibrio in the water sample and shrimp samples at the initial and the end of the culture period in the challenge test when treated with control commercial feed (CV), prebiotic feed (PV), and synbiotic feed (S9V).
4 Discussion
4.1 Proximate analysis
The addition of H. alkaliphila was found to increase the carbohydrate content in the synbiotic feed. According to Aquacop (1978), bacteria can contribute to the bulk of carbohydrate, especially when shrimp are reared in an earthen pond or super-intensive conditions. Additionally, the K. alvarezii contains polysaccharides that also can increase the feed carbohydrate content (Suantika et al., 2017).
4.2 Water quality in the shrimp culture system
During performance and challenges test, the physicochemical properties of the culture system; including dissolved oxygen, pH, temperature, salinity, as well as ammonium, nitrite, and nitrate levels remain in the optimum range for shrimp cultivation. Hence, the water quality has no negative effect on the survival and growth of L. vannameii.
4.3 Biological parameters of L. vannameii after the feeding and challenge tests
Synbiotic feed supplementation resulted in beneficial effects for L. vannamei post-larval cultures in this study. The higher survival and vibriosis resistance even after the challenge test against V. harveyii indicates that the probiotics and prebiotics in the tested synbiotics acted synergistically. Hypothetically, the mechanism behind this effect is complex: first, K. alvarezii is used as a nutritional source for probiotic bacteria, which enhances their growth while limiting that of opportunistic and potentially pathogenic bacteria. Then, the halophilic H. alkaliphila produces the enzyme alpha-glucosidase (Romano et al., 2006), which suggests that H. alkaliphila can potentially hydrolyze oligosaccharides and polysaccharides (non-digestible carbohydrates) in K. alvarezii. The metabolic results of non-digestible carbohydrates can be in the form of short-chain fatty acids (SCFA), which can stimulate the activation of the immune system (Morrison and Preston, 2016). Halomonas sp. are also through to cause Vibrio quorum quenching. The lactonase produced by Halomonas sp. can hydrolyze N-acylhomoserine lactone (AHL) which is an autoinducer in Vibrio quorum sensing (Romano et al., 2006; Fangfang et al., 2008).
Vibrio spp. Are natural opportunistic micro-flora of shrimps (Sindermann, 1990) and may become pathogenic when shrimps are in sub-optimal condition, such as physiological unbalance and nutritional deficiency, or when there are other stressors, i.e., poor water quality and overstocking, which allow opportunistic bacterial infections to proress (Brock and Lightner, 1990). Indeed, inappropriate levels of shrimp larval density and poor living conditions of shrimp larvae may also cause Vibrio to become pathogenic (Brock and Lightner, 1990). The shrimp density used in this study was very high compared to the common shrimp stocking density of 60–300 PL.m−3 (FAO, 2009; Suantika et al., 2018a,b,c, 2020). In normal conditions, high stocking density may affect shrimp survival, because the Vibrio population can be pathogenic in the batch shrimp culture system. This might in turn result in high shrimp mortality (up to 70%), even in treatment groups without Vibrio addition. However, the use of synbiotic feed still provided the highest shrimp survival, suggesting it also increased vibriosis resistance.
4.4 Bacterial community during feeding test and challenge test
The use of synbiotic feed is known to induce the growth of beneficial intestinal bacteria (Pandey et al., 2015). In this study, P. piscicida and B. cereus isolated from shrimp and culture water samples subjected to the synbiotic treatment could be used as a probiotic to increase disease resistance (Far et al., 2009; Ramadhani et al., 2019). Vibrio quorum quenching in shrimp and culture water can be possible due to synbiotic-feed treatment, so the pathogenicity of Vibrio will decrease. Lactonase may have produced by H. alkaliphila which can hydrolyze AHL (N-acylhomoserine lactone) as autoinducer in Vibrio quorum sensing (Romano et al., 2006; Fangfang et al., 2008). In this study, the increased of beneficial bacteria in synbiotic treatment may also reduce the dominance of Vibrio.
5 Conclusion
In conclusion, supplementing the commercial feed used in shrimp aquaculture with K. alvarezii and Spirulina sp. prebiotics and H. alkaliphila probiotic enhance the survival of L. vannamei in post-larval cultures, under both normal conditions and when infected with V. harveyii. This suggests that the tested synbiotic feed can potentially be used as a vibriosis biocontrol agent in intensive shrimp aquaculture.
Acknowledgements
This research was funded by Ministry of Research, Technology, and Higher Education of the Republic of Indonesia under the program “Penelitian Terapan Unggulan Perguruan Tinggi” 2018–2019.
References
- Anuta JD, Buentello A, Patnaik S, Hume ME, Mustafa A, Gatlin DM, Lawrence AL. 2014. Effects of dietary supplementation of a commercial prebiotic previda® on survival, growth, immune reponses and gut microbiota of pacific white shrimp, Litopenaeus vannamei Aquac Nutr 1: 1–9. [Google Scholar]
- Aquacop. 1978. Study of nutritional requirements and growth of Penaues merguiensis in tanks by means of purified and artificial diets. Proc Maricult Soc 9: 225–234. [Google Scholar]
- Arisa II, Widanarni W, Yuhana M, Muchlisin ZA, Muhammadar AA. 2015. The application of probiotics, prebiotics, and synbiotics to enhance the immune responses of vannamei shrimp (Litopenaeus vannamei) to Vibrio harveyi Infection. AACL Bioflux 8: 772–778. [Google Scholar]
- Boyd CE, Nevin AAM. Aquaculture Resource Use and the Environment, Wiley-Blackwell, New Jersey USA, 2015. [Google Scholar]
- Brock JA, Lightner DV. 1990. Diseases of Crustacea. Diseases caused by microorganisms, in: O. Kinne (ed.). Diseases of Marine Animals. Volume III: Introduction, Cephalopoda, Annelida, Crustacea, Chaetognatha, Echinodermata, Urochordata, Biologische Anstalt Helgoland, Hamburg, pp. 309–318. [Google Scholar]
- Cappuccino JG, Sherman N. 2014. Microbiology: A Laboratory Manual, 10th edn, Pearson, USA. [Google Scholar]
- Cencic A, Chingwaru W. 2010. The role of functional foods, nutraceuticals, and food supplements in intestinal health, Nutrients 2: 611–625. [PubMed] [Google Scholar]
- Chandran N, Iyapparaj P, Moovendhan S, Ramasubburayan R, Prakash S, Immanuel G, Palavesam A. 2014. Influence of probiotic bacterium Bacillus cereus isolated from the intestine of wild shrimp Penaeus monodon in turn as potent growth promoter and immune enhancer in P. monodon, Fish Shellfish Immunol 36: 38–45. [PubMed] [Google Scholar]
- Chiu C, Guu Y, Liu C. 2007. Immune responses and gene expression in white shrimp Litopenaeus vannamei induced by Lactobacillus plantarum, Fish Shellfish Immunol 23: 364–377. [PubMed] [Google Scholar]
- Das S, Mondal K, Haque S. 2017. A review on application of probiotic, prebiotic and synbiotic for sustainable development of aquaculture, J Entomol Zool Stud 5: 422–429. [Google Scholar]
- Davis DA. Nutrition Requirements of Penaid Shrimp, Auburn University, Auburn, Alabama, USA, 2005. [Google Scholar]
- Fangfang B, Yin H, Jixiang C, Xiao-Hua Z. 2008. Disruption of quorum sensing in Vibrio harveyi by the AiiA protein of Bacillus thuringiensis, Aquaculture 1: 36–40. [Google Scholar]
- FAO. Penaeus vannamei, in: Cultured Aquatic Species Fact Sheets, Food and Agriculture Organization of the United Nations, Rome, 2009. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture: Opportunities and Challenge, Food and Agriculture Organization of the United Nations, Rome, 2014. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture: Opportunities and Challenges, Food and Agriculture Organization of the United Nations, Rome, 2018. [Google Scholar]
- FAO. Globefish Highlights A Quarterly Update on World Seafood Market, Food and Agriculture Organization of the United Nations. Rome, 2019. [Google Scholar]
- Far HZ, Saad CRB, Daud HM, Harmin SA, Shakibazadeh S. 2009. Effect of Bacillus subtilis on the Growth and Survival Rate of Shrimp (Litopenaeus vannamei). Afr J Biotechnol 8: 3369–3376. [Google Scholar]
- Glencross BD, Huyben D, Schrama JW. 2020. The application of single-cell ingredients in aquaculture feeds − a review, Fishes 5(22): 1–39. [Google Scholar]
- Hardjani D, Suantika G, Aditiawati P. 2017. Nutritional profile of red seaweed Kappaphycus alvarezii after fermentation using Saccharomyces cerevisiae as a feed supplement for white shrimp Litopenaeus vannamei nutritional profile of fermented red seaweed, J Pure Appl Microbiol 11: 1637–1645. [Google Scholar]
- Huynh TG, Cheng AC, Chi CC, Chiu KH, Liu CH. 2018. A synbiotic improves the immunity of white shrimp, Litopenaeus vannamei: metabolomic analysis reveal compelling evidence, Fish Shellfish Immunol 79: 284–293. [PubMed] [Google Scholar]
- Ibrahem MD, Mohamed MF, Ibrahim MA. 2013. The role of Spirulina platensis in growth and immunity of nile tilapia and its resistance to bacterial infection, J Agric Sci 5: 109–117. [Google Scholar]
- Jalali M, Abedi D, Varshoaz J, Najjarzadeh M, Mirlohi M, Tavakoli N. 2012. Stability evaluation of freeze-dried Lactobacillus paracasei subsp. tolerance and Lactobacillus delbrueckii subsp. bulgaricus in oral capsules, Res Pharm Sci 7: 31–36. [PubMed] [Google Scholar]
- Kechagia M, Basoulis D, Konstantopoulou S, Dimitriadi D, Syftopoulou K, Skarmoutsou N, Fakiri EM. 2013. Health benefits of probiotics: a review, ISRN Nutr 1: 1–7. [Google Scholar]
- Lemos LN, Fulthorpe RR, Triplett EW, Roesch LF. 2011. Rethinking microbial diversity analysis in the high throughput sequencing era. J Microbiol Methods 86: 42–51. [PubMed] [Google Scholar]
- Maehre HK, Dalheim L, Edvinsen GK, Elvevoll EO, Jensen IJ. 2018. Protein determination- method matters, Foods 7: 5 [Google Scholar]
- Morrison DJ, Preston T. 2016. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 7: 189–200. [PubMed] [Google Scholar]
- Oktaviana A, Widanarni, Yuhana M. 2014. The use of synbiotics to prevent IMNV and Vibrio harveyi Co-infection in Litopenaeus vannamei, HAYATI J Biosci 21: 127–134. [Google Scholar]
- Pandey KR, Naik SR, Vakii BV. 2015. Probiotics, prebiotics, and synbiotics − a review, J Food Sci Technol 52: 7577–7587. [PubMed] [Google Scholar]
- Ramadhani DE, Widanarni W, Sukenda S. 2019. Microencapsulation of probiotics and its applications with prebiotic in Pacific white shrimp larvae through Artemia sp., JAI 18: 130–140. [Google Scholar]
- Ramezani FE, Zokaeifar H, Ebrahimi M, Kamarudin MS, Goh YM, Ehteshami F. 2014. Probiotic administration of Litopenaeus vannamei: is there any negative effect on the fatty acid profile of meat, Iran J Fish Sci 13: 550–559. [Google Scholar]
- Romano I, Lama L, Nicolaus B, Poli A, Gambacorta A, Giordano A. 2006. Halomonas alkaliphila sp nov., a novel halotolerant alkaliphilic bacterium isolated from a salt pool in Campania (Italy). J Gen Appl Microbiol 52: 339–348. [PubMed] [Google Scholar]
- Savani M, Cecchini C, Verdenelli MC, Silvi S, Orpianesi C, Cresci A. 2010. Pilot-scale production and viability analysis of freeze-dried probiotic bacteria using different protective agents, Nutrients 2: 330–339. [PubMed] [Google Scholar]
- Sindermann CJ. 1990. Principal Diseases of Marine Fish and Shellfish, Vol. 2, 2nd edition. Academic Press, New York. [Google Scholar]
- Situmorang ML, Suantika G, Santoso M, Khakim A, Wibowo I, Aditiawati P, Haniswita. 2020. Poly‐β‐Hydroxybutyrate (PHB) improves nursery‐phase pacific white shrimp Litopenaeus vannamei defense against vibriosis, N Am J Aquac 82: 108–114. [Google Scholar]
- Smith P. 2008. Antimicrobial resistance in aquaculture, Rev Sci Tech 27: 243–264. [PubMed] [Google Scholar]
- Suantika G, Aditiawati P, Astuti DI, dan Khotimah ZF. 2013. The use of indigenous probiotic Halomonas aquamarina and Shewanella algae for white shrimp (Litopenaeus vannamei Boone) hatchery productivity in Zero Water Discharge system. J Aquac Res Dev 4: 1–8. [Google Scholar]
- Suantika G, Situmorang ML, Aditiawati, Khakim A, Suryanarayan S, Nori SS, Kumar S, Putri F. 2017. Effect of red seaweed Kappaphycus alvarezii on growth, salinity stress tolerance and vibriosis resistance in shrimp Litopenaeus vannamei hatchery, J Fish Aquatic Sci 12: 127–133. [Google Scholar]
- Suantika G, Situmorang ML, Nurfathurahmi A, Taufik I, Aditiawati P, Yusuf N, Aulia R. 2018a. Application of indoor recirculation aquaculture system for white shrimp (Litopenaeus vannamei) grow-out super-intensive culture at low salinity condition, J Aquac Res Dev 9: 1–6. [Google Scholar]
- Suantika G, Situmorang ML, Khakim A, Wibowo I, Aditiawati P, Suryanarayan S, Nori SS, Kumar S, Ferisca P. 2018b. Effect of red seaweed Kappaphycus alvarezii on growth, survival, and disease resistance of pacific white shrimp Litopenaeus vannamei against Vibrio harveyi in the nursery phase, J Aquac Res Dev 9: 1–7. [Google Scholar]
- Suantika G, Situmorang ML, Kurniawan JB, Pratiwi SA. 2018c. Development of a Zero Water (ZWD) − Recirculating Aquaculture System (RAS) hybrid system for super intensive white shrimp (Litopenaeus vannamei) culture under low salinity conditions and its industrial trial in commercial shrimp urban farming in Gresik, East Java, Indonesia, Aquac Eng 82: 12–24. [Google Scholar]
- Suantika G, Situmorang ML, Saputra FI, Putri SLE, Putri SP, Aditiawati P, Fukusaki E. 2020. Metabolite profiling of whiteleg shrimp Litopenaeus vannamei from super‑intensive culture in closed aquaculture systems: a recirculating aquaculture system and a hybrid zero water discharge–recirculating aquaculture system, Metabolomics 16: 49. [PubMed] [Google Scholar]
- Zhang L, Kangsen M, Tan B, Qinghui A, Chiengzhien Q. 2009. Effect of dietary administration of probiotic Halomonas sp. B12 on the intestinal microflora, immunological parameters and midgut histological structure of shrimp, Fenneropenaeus chinensis, J World Aquac Soc 40: 58–66. [Google Scholar]
- Zhou X, Wang Y, Li W. 2009. Effect of probiotic on larvae shrimp (Penaeus vannamei) based on water quality, survival rate, and digestive enzyme activities, Aquaculture 287: 349–353. [Google Scholar]
Cite this article as: Situmorang ML, Nurwidayanti P, Suantika G. 2021. Synbiotic containing Kappaphycus alvarezii, Spirulina sp., and Halomonas alkaliphila improves survival, growth, and vibriosis resistance in whiteleg shrimp (Litopenaeus vannamei) post-larval culture. Aquat. Living Resour. 34: 10
All Tables
Proximate analysis results of commercial feed (C), prebiotic-supplemented feed (P), synbiotic-supplemented feed using Halomonas alkaliphila 108 CFU.kg−1 (S8), Halomonas. alkaliphila 109 CFU.kg−1 (S9), and Halomonas alkaliphila 109 CFU.kg−1 (S10) used in this experiment, with the optimum range values provided for comparison.
The effect of the 14-day feeding test on Litopenaeus. vannamei using control commercial feed (C); prebiotic-supplemented feed (P); and synbiotic-supplemented feed with Halomonas alkaliphila 108 CFU.kg−1 (S8), 109 CFU.kg−1 (S9), and 1010 CFU.kg−1 (S10).
Results of the 7-day challenge test against Vibrio harveyi on post-larval Litopenaeus vannamei with control commercial feed (C), prebiotic-supplemented feed (P), and synbiotic-supplemented feed with Halomonas alkaliphila 109 CFU.kg−1 (S9).
The total number of Vibrio in the water sample and shrimp samples at the initial and the end of the culture period in the challenge test when treated with control commercial feed (CV), prebiotic feed (PV), and synbiotic feed (S9V).
All Figures
|
Fig. 1 The Shannon-Weaver index of the bacterial community in shrimp culture and water culture water during the feeding test at day 0 (T0), day 7 (T1), and day 14 (T2) during the feeding test using commercial feed (C), prebiotic-supplemented feed (P), synbiotic-supplemented feed using Halomonas alkaliphila 108 CFU.kg−1 (S8), Halomonas alkaliphila 109 CFU.kg−1 (S9), and Halomonas alkaliphila 109 CFU.kg−1 (S10). |
| In the text | |
|
Fig. 2 Percentage of dominant bacterial community in shrimp culture (a) and water culture (b) at day 0 (T0), day 7 (T1), and day 14 (T2) during the feeding test using commercial feed (C), prebiotic-supplemented feed (P), synbiotic-supplemented feed using Halomonas alkaliphila 108 CFU.kg−1 (S8), Halomonas alkaliphila 109 CFU.kg−1 (S9), and Halomonas alkaliphila 109 CFU.kg−1 (S10). |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.