| Issue |
Aquat. Living Resour.
Volume 33, 2020
|
|
|---|---|---|
| Article Number | 18 | |
| Number of page(s) | 11 | |
| DOI | https://doi.org/10.1051/alr/2020018 | |
| Published online | 30 November 2020 | |
Research Article
Growth, mortality and yield of Sarotherodon melanotheron melanotheron (Rüppell, 1852) in the Lake Nokoué and Porto-Novo Lagoon complex Benin, West Africa
1
Laboratory of Hydrobiology and Aquaculture, Faculty of Agricultural Sciences, University of Abomey-Calavi, Cotonou, Benin
2
Laboratory of Animal Pathology, Microbiology and Immunology, University of Abomey-Calavi, Cotonou, Benin
3
Department of Aquatic Environment, INP/ENSAT, Castanet Tolosan, France
* Corresponding author: This email address is being protected from spambots. You need JavaScript enabled to view it.
Handling Editor: Verena Trenkel
Received:
20
July
2020
Accepted:
22
October
2020
Abstract
The study evaluated some population parameters of Sarotherodon melanotheron melanotheron within a lagoon complex in southern Benin: Lake Nokoué and Porto-Novo Lagoon. Data on the total length, total weight, and sex were recorded monthly between January and December 2015 for 1,745 specimens captured by local fishermen. The asymptotic length L∞ was estimated at 24.68 cm. The growth rate K was 0.86. The total, natural, and fishing mortalities were estimated at 2.46, 1.71, and 0.75 y−1, respectively. The size at first sexual maturity was 8.9 and 9.2 cm, respectively, for males andfemales. The size at first capture was estimated at 9.2 cm, which indicates that fish spawn at least once before capture. The current exploitation rate of 0.31 remains below the maximum exploitation rate Emax which was 0.604 but was equal to the exploitation rate retaining 50% of the biomass of the stock (E0.5 = 0.324). This suggests that the stock of S. m. melanotheron is not overexploited in the complex. However, it is recommended that the actual rate of exploitation be kept as it is to ensure a sustainable management of these fish populations.
Key words: Cichlid / ELEFAN / Lagoon fisheries / Ouémé river / Demographic parameters
© EDP Sciences 2020
1 Introduction
In Benin, several studies meant to update data on fish fauna inventory and its exploitation have been carried out in recent years (Lalèyè, 1995; Lalèyè et al., 2003, 2004; Moritz et al., 2006; Montchowui et al., 2007; Moritz, 2010; Ahouansou Montcho, 2011; Lederoun et al., 2015, 2016, 2018a,b; Adjibade et al., 2019; Imorou et al., 2019; Moritz and Lalèyè, 2019) but those studies have mostly focused on large river basins, leaving lakes and lagoons often heavily exploited by the local population (Lederoun et al., 2015, 2016). In southern Benin, there are some lakes and lagoons along the coastline: Lake Ahémé, the Coastal lagoon, the old lagoons (Toho, Todougba, Dati, Ahoungan, and Djonou), and the Lake Nokoué and Porto-Novo Lagoon complex. This lagoon complex remains the most important waterbody from the point of view of its extent, productivity, and exploitation (Lalèyè, 2000). Overall, this latter component, i.e., exploitation, has been very little-documented in recent years despite fishing pressure on the target species (Lalèyè, 2000). Indeed, the latest available information on the exploitation of the complex's resources is that of Niyonkuru (2007) on Lake Nokoué, highlighting the heavy massive exploitation of three cichlids (Sarotherodon melanotheron, Coptodon guineensis, and Hemichromis fasciatus). Since then, changes have been noted in this ecosystem and especially in the entire lagoon system. In fact, over the past decade, the aquatic space of the complex has been taken over by man for various uses. This was mainly because of anarchic occupation of the two bodies of water by the fishermen for the establishment of sedentary fisheries through acadjas (a type of fish parks composed either of branches stuck in the bottom of water or of floating vegetation), thus reducing the aquatic space available for other fishing practices (Vodougnon et al., 2018).
Available national data of 1998 show that inland fishing produces 33,000 tonnes y−1, 6,000 tonnes of which come from the acadjas (Aglinglo, 1998). Most of these catches are provided by brackish water bodies (95% of the total, i.e., 31,000 tonnes). Lake Nokoué alone accounts for more than 69% of the total production (22,000 tonnes). According to statistics from the Fisheries Production Department, the production of Lake Nokoué increased from 1993 to 1996, going from 18,000 tonnes to 21,900 tonnes. This increase is mainly due to the rise in the number of acadjas, which went from 589 (covering 311.8 ha) in 1981 to 9,078 (5,931 ha) in 1996 (Aglinglo, 1998). Even if updated statistics do not currently exist, the number of acadjas and space occupied by these sedentary fisheries has increased considerably in recent years, impacting the number of fishermen. We have gone from nearly 17,000 fishermen in 1990s–96 to 11,492 from 1997. The decrease in the number of fishermen from almost 17,000 in 1990s–96 to 11,492 in 1997 in favor of other activities is a sign of reduced yield despite increased fishing effort. However, in addition to this very controversial method of exploitation, there is also the use of devastating fishing gear and techniques in the few open spaces, such as small mesh nets, which are often reported in the complex. Faced with all these disturbances and threats mentioned on these fish species, there is an urgent need to update data on the most exploited fish species' population dynamics in the Lake Nokoué and Porto-Novo Lagoon complex. Among these most exploited species are the Cichlidae, Sarotherodon melanotheron melanotheron. Indeed, statistical surveys (Lalèyè et al., 2003; Niyonkuru, 2007) have shown that S. m. melanotheron is most abundant in Lake Nokoué with Coptodon guineensis (Cichlidae) and Ethmalosa fimbriata (Clupeidae). Overall, S. m. melanotheron represents 72% of the total catches of fish of the Cichlidae family in Lake Nokoué outside the acadjas and nearly 87% in the acadjas (Niyonkuru, 2007). A recent study on the exploitation of the species in Lake Toho in Benin (Lederoun et al., 2015) showed that S. m. melanotheron has reached high levels of exploitation due to increasing demand in recent years, and its economic value. In the Lake Nokoué and Porto-Novo Lagoon complex, where the species is also abundant, up-to-date information on demographic parameters is lacking, which significantly limits decision-making in terms of the management of renewable resources exploited.
This is the context within which this study is conducted, aiming to assess the population dynamics and exploitation of Sarotherodon melanotheron melanotheron, one of the main species caught by fishermen in the Lake Nokoué and Porto-Novo Lagoon complex.
2 Materials and methods
2.1 Study area
The fish were caught in Lake Nokoué (6°20′ to 6°30′N and 2°20′ to 2°35′E) and the Porto-Novo Lagoon (6°25′ to 6°30′N and 2°30′ to 2°38′E) in Benin (Fig. 1). Lake Nokoué, which connects to the Porto-Novo Lagoon through the Totchè canal, covers an area of about 150 km2 and is freshwater-fed by both the Ouémé and Sô Rivers (Niyonkuru and Lalèyè, 2010; Vodougnon et al., 2018). It is also affected by the Cotonou tidal channel through which seawater reaches it (Lalèyè, 1995; Lalèyè et al., 2003; Vodougnon et al., 2018). Less stretched than the Nokoué Lake, the Porto-Novo Lagoon covers 35 km2 (Lalèyè, 1995; Gnohossou, 2006). It is the point through which the Ouémé River's waters reach the ocean through the Lagos tidal channel. Lake Nokoué and Porto-Novo Lagoon complex are subject to a sub-equatorial climate characterized by an alternation of two rainy seasons (early April through mid-July and mid-September through mid-November) and two dry seasons (mid-November through late March and mid-July through mid-September) due to its geographical location (Vodougnon et al., 2018).
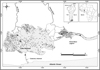 |
Fig. 1 Study area, showing sampling sites (▲) and acadjas (■). |
2.2 Sampling methods
Between January and December 2015, fish samples (N = 1,745) were collected randomly from artisanal fishermen who operated scoop nets, monofilament gillnets, casts nets, and acadjas at six sampling sites (Fig. 1). They were transported to the laboratory for identification using the key by Teugels and Thys van den Audenaerde (2003). Each specimen was measured for total length to the nearest 0.1 cm using measuring-tape, individually weighed (fresh weight) to the nearest 0.01 g using an electronic precision balance, and sexed (immature, male, or female) following the criteria of Brown-Peterson et al. (2011). Some of the fish were deposited in the Royal Museum for Central Africa (RMCA), Tervuren, Belgium, to establish a reference collection.
2.3 Data processing
The relationship between total length and the total weight was established using the equation: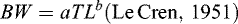 (1)
(1)
where BW is wet body weight (g), TL total length (cm), a the intercept, and b the slope of the linear regression. The 95% confidence limits for b were assessed using Statview software, version 1992–98 (SAS Institute INC). In order to check whether b is significantly different from 3, the Student t-test was conducted following Sokal and Rohlf (1987):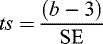 (2)where ts is the t-test value, b the slope, and SE the standard error of b.
(2)where ts is the t-test value, b the slope, and SE the standard error of b.
Size at first maturity (L50) was estimated for both sexes using a logistic function that was fitted to the proportion of sexually mature individuals by each size class using a non-linear regression following King's (1995) formula: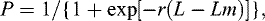 (3)
(3)
where P is the proportion mature in each size class, r (−b slope) is a parameter controlling the slope of the curve, and Lm is the size at which 50% of fish are mature (Saila et al., 1988). Statistical analyses and plots were generated using R (R Development Core Team, 2018) with sizeMat (Torrejon-Magallanes, 2019) package. Likelihood-ratio tests were performed to detect differences in L50 values between sexes.
For the growth analysis, the length-frequency data were pooled into groups with 1 cm length class intervals. The growth parameters such as asymptotic length (L∞) and growth rate (K) were obtained using Von Bertalanffy Growth Function (VBGF). The ELEFAN I (electronic length-frequency analysis) software tool included in FiSAT II (FAO ICLARM Stock Assessment Tools) (Gayanilo et al., 1996) was used to estimate the parameters L∞ and K of the von Bertalanffy equation. According to VBGF, individual fishes grow on average towards the asymptotic length at an instantaneous growth rate (K) with length on time (t) following the expression: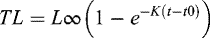 (4)where TL is the total length of the fish at time t, L∞ is the asymptotic length of fish (cm), K is the rate at which TL approaches L∞, and t0 is the theoretical age of the fish when TL is equal to zero.
(4)where TL is the total length of the fish at time t, L∞ is the asymptotic length of fish (cm), K is the rate at which TL approaches L∞, and t0 is the theoretical age of the fish when TL is equal to zero.
The theoretical age at length zero (to) was estimated by the equation of Pauly (1979):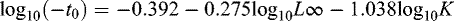 (5)
(5)
The longevity, tmax, was calculated using the following formula (Taylor, 1962; Pauly, 1980):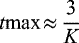 (6)
(6)
The growth performance index (phi prime, Φ′) was estimated using the equation of Pauly and Munro (1984):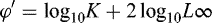 (7)
(7)
The total mortality coefficient Z (y−1) was estimated by a length-converted catch curve (Pauly, 1984).
The natural mortality rate M (y−1) which caused by all other factors except fishing, was estimated according to Pauly's (1980) empirical formula: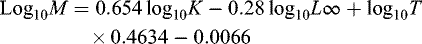 (8)
(8)
where K and L∞ are the parameters of the equation (4) and T the average annual temperature of the water which is 28 °C. The Fishing mortality rate (F) was estimated using the relation: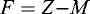 (9)
(9)
The exploitation rate (E) was calculated by the quotient between fishing and total mortality (Pauly, 1984):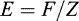 (10)
(10)
The length at which 50% of the fishes are selected by the gear (Lc = size at first capture) was estimated using the ogive selection (Pauly, 1984).
The optimal length Lopt was estimated using Froese's (2004) equation: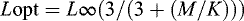 (11)where L∞ and K are function of the von Bertalanffy growth and M the natural mortality rate.
(11)where L∞ and K are function of the von Bertalanffy growth and M the natural mortality rate.
Relative yield per recruit (Y ′/R) was estimated using Beverton and Holt's (1966) model as follows: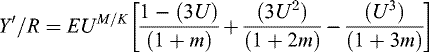 (12)where E = F/Z corresponds to the current exploitation rate, i.e. the fraction of the total mortality caused by fishing activities. U = 1−(Lc/L∞) = the fraction of growth to be completed by the fish after its entry into the exploitation phase (Lc = mean length at first capture, L∞ = asymptotic length), m = (1−E)/(M/K) = K/Z. The Relative Y ′/R analysis assumed that the fish species conforms to knife-edge recruitment and knife-edge selection. The relative biomass per recruit (B ′/R) was estimated as:
(12)where E = F/Z corresponds to the current exploitation rate, i.e. the fraction of the total mortality caused by fishing activities. U = 1−(Lc/L∞) = the fraction of growth to be completed by the fish after its entry into the exploitation phase (Lc = mean length at first capture, L∞ = asymptotic length), m = (1−E)/(M/K) = K/Z. The Relative Y ′/R analysis assumed that the fish species conforms to knife-edge recruitment and knife-edge selection. The relative biomass per recruit (B ′/R) was estimated as: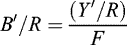 (13)
(13)
The recruitment patterns were computed following the method described in the FiSAT routine (Gayalino et al., 1996).
3 Results
3.1 Size frequencies and length-weight relationship
For 1,745 S. m. melanotheron samples collected between January and December 2015, the total length ranged from 4.6 to 23.1 cm, with an average of 11.6 ± 3.3 cm (Fig. 2). The weight of caught fishes varied between 2.76 and 286.43 g with an average of 36.44 ± 30.25 g. The length-weight relationships was described by the equation W = 0.036 TL2.742; (r 2 = 0.963; SE = 0.137) with a strong correlation (see r 2) linked here to the wide size range of sampled fish (Fig. 2), the difference between the extreme length values being 18.5 cm. The allometric coefficient b was significantly lower than 3 (ts = −1.88; confidence interval at 95% for b: 2.715–2.769), so growth was a negative allometric growth.
 |
Fig. 2 Size frequencies for Sarotherodon melanotheron melanotheron (Cichlidae) in Lake Nokoué and Porto-Novo Lagoon complex (N = 1,745). |
3.2 Size at first maturity
The logistic fit method suggests 50% of female S. m. melanotheron in the Lake Nokoué and Porto-Novo Lagoon complex are sexually mature at 9.2 cm TL with 95% confidence intervals ranging from 9.1 to 9.4 cm TL (Fig. 3). For males, corresponding values are 8.9 cm TL et 8.5–9.2 cm TL for size at first maturity and confidence intervals, respectively (Fig. 3). There was no significant difference in the estimated size at maturity between males and females (p = 0.754).
 |
Fig. 3 Estimated size at first maturity for males (a) and females (b) of Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex. |
3.3 Growth parameters
The asymptotic length L∞ and the growth coefficient K were 24.68 cm and 0.86 y−1, respectively. The growth performance index Φ′ was relatively high for this species, estimated at 2.42, and the theoretical ages at which length is 0 was estimated at −0.2 year. The potential longevity tmax was 3.49 years. Specimens of S. m. melanotheron captured during the study period belonged to 4 cohorts (Fig. 4).
 |
Fig. 4 Length frequency data superimposed on the growth curve (Continuous line) of Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex (L∞ = 24.68 cm total length, K = 0.86 y−1). The bars represent the restructured length frequency data, where black bars indicate positive peaks and white bars represent negative peaks. |
3.4 Mortality and exploitation rate
The total mortality (Z) estimate from the length converted catch curve was 2.46 y−1 (Supplemental Fig. S1). The natural mortality rate (M) and fishing mortality rate (F) were 1.71 y−1 and 0.75 y−1, respectively. The current exploitation rate was estimated as E = 0.31.
3.5 Size at first capture and optimal size
The length at first capture (Lc), i.e., the length at which 50% of the S. m. melanotheron species are vulnerable to capture, was estimated at 9.2 cm (Fig. 5). The logistics model selection also showed that 25% of fish at 6.6 cm and 75% of fish at 11.9 cm were caught. All fish over 23.5 cm were systematically caught. The estimated optimal length was 14.8 cm.
 |
Fig. 5 Probability of capture of Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex estimated from the ascending axis of the catch curve. |
3.6 Relative yield per recruitY ′/ R and reference points
In the Lake Nokoué and Porto-Novo Lagoon complex, the relative yield curves per recruit Y ′ /R relative to the exploitation ratio E, indicated an optimal exploitation rate (Emax) of 0.610 (Fig. 6). The exploitation rate E0.1 (exploitation rate at which the marginal increase of Y ′ /R is 10% of its entire stock) and E0.5 (exploitation rate under which the whole stock is halved) were respectively estimated at 0.515 and 0.325.
 |
Fig. 6 Relative yield-per-recruit and biomass-per-recruit curves for Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex using the selection ogive option. The three dashed right-angled lines correspond to E0.5, E0.1 and Emax, respectively. |
3.7 Recruitment patterns
Recruitment patterns of S. m. melanotheron from the Lake Nokoué and Porto-Novo Lagoon suggested that recruitment was continuous throughout the year with two distinct spawning events (minor and major) (Fig. 7). The minor peak of recruitment occurred in March-April with a recruitment strength of 12.60% and 12.61%, respectively, whereas the significant rise occurred in August with a recruitment strength of 18.96%.
 |
Fig. 7 Recruitment pattern of Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex showing two recruitment peaks per year. |
4 Discussion
Length-weight relationships play a significant role in assessing the biology of fish fauna (Abowei et al., 2009; Akintola et al., 2010). The relationships between length and weight (with coefficient b = 2.742) for 1,745 specimens of S. m. melanotheron indicates a negative allometric growth. Which means that, the increase in length was faster than the growth in weight in this study. This result is consistent with that of Lederoun et al. (2015), who reported negative allometric growth (b = 2.9593) for this species in Lake Toho in Benin. The same kind of growth (b = 2.812) has also been reported for the same species in the Ayamé I reservoir in Ivory Coast (Tah et al., 2012) and from the coastal rivers (b = 2.815) of the south-eastern part of Ivory Coast (Konan et al., 2007).
The size of 8.9 and 9.2 cm TL first sexual maturity obtained respectively for males and females is more significant than that reported by Lederoun et al. (2015) for the same species in Lake Toho, i.e., 6.9 and 7.7 cm TL for males and females, respectively. Apart from the fishing pressure reported in Lake Toho by Lederoun et al. (2015), the presence in number and on large surfaces of acadjas in Lake Nokoué (1933 acadjas, 22.13 km2) and Porto-Novo Lagoon (474 acadjas, 7.93 km2) (see Fig. 1) complex could explain this size difference. Indeed, acadja is a vast artificial fish aggregation device made up of clumps of branches implanted in shallow areas of a water body. The installation generally allows the fish to shelter from predators and feed on the microorganisms that colonize this environment, which would increase the size of the fish and catch per unit area compared to open water fishing. The hiding places constituted by the acadjas would also help reduce the fishing pressure on the fish before their exploitation, which generally takes place at least one year after their installation. It is a traditional fishing practice that is profitable if well managed (Welcomme, 1972; Lalèyè, 2000). The size at first maturity for S. m. melanotheron in the complex remains lower than that obtained in Lake Doukon (12.8 and 13.2 cm for males and females, respectively) and Lake Togbadji (12.4 and 11.5 for males and females, respectively) for Sarotherodon galilaeus galilaeus (Lederoun et al., 2016). The reduction in size at first sexual maturity as a function of fishing pressure is well known among cichlids (Leonardis and Sinis, 1998; Panfili et al., 2004; Lederoun et al., 2015).
According to Alhassan and Armah (2011), estimation of growth and mortality is fundamental in fisheries because stock assessment and management rely on these population parameters. There are several mathematical models for expressing change. Von Bertalanffy's model remains the most used (Hilborn and Walters, 1992). The von Bertalanffy model's growth parameters are often obtained using size frequency distributions, especially for tropical fish, to circumvent the difficulties linked to the interpretation of growth marks on hard parts (otolithometry, e.g.). In the present study, this model has been used to estimate growth parameters and those of mortalities. The model has the advantage of accurately describing the growth in the interval of the observed data, and it is easily incorporated into production models (Gaamour, 1999; Rjeibi et al., 2011). In principle, statistical software MIX (MacDonald and Pitcher, 1979), MULTIFAN (Tuck et al., 1997), and ELEFAN are excellent examples (Wahle and Fogarty, 2006). Still, in this study, ELEFAN, designated by ELEFAN I in the FiSAT II fisheries analysis software, was chosen. ELEFAN allows an estimation of growth parameters by a direct projection of frequency-size data without first translating the length scale into an age scale (Gayanilo et al., 2005). ELEFAN has been widely used in previous studies on Lake Nokoué (Villanueva, 2004; Niyonkuru, 2007; Niyonkuru et al., 2007) and elsewhere (King and Etim, 2004; Montchowui et al., 2009; Montchowui et al., 2011; Lederoun et al., 2015, 2016) and therefore remains the most appropriate in cases of growth comparison between populations and species.
In this study, the growth coefficient K was estimated at 0.86 y−1; the asymptotic length L∞ obtained was 24.68. Referring to the criteria for determining the modes of growth of Branstetter (1987) who estimated that in a given species, growth is slow when 0.05 < K <0.10 y−1, intermediate when 0.10 < K <0.20 y−1, rapid when 0.20 < K <0.50 y−1, and very rapid when K ≥ 0.5, S. m. melanotheron has a very rapid growth in the Lake Nokoué and Porto-Novo Lagoon complex, which agrees with the good primary productivity of the complex (Sarmento et al., 2009). The four generations identified in the complex are lower than the six generations identified in Lake Toho (Lederoun et al., 2015) for the same species. Different environmental conditions could explain this situation. The relatively high fishing pressure in Lake Toho would favor rapid multiplication to the disadvantage of growth.
The estimated growth performance index (ϕ ′ = 2.42) is similar to those found in the literature (Tab. 1). The slight variations observed would be due to the availability of food, to population density (Lederoun et al., 2015, 2016; Bédia et al., 2017), differences in accessibility to food (Pauly, 1994), differences in ecosystem temperatures (King and Etim, 2004), environmental pollution, fisheries degradation (Abowei and Hart, 2007, 2009) and fishing pressure (Lederoun et al., 2015).
March-April and August periods are those of great recruitments for S. m. melanotheron in the study site. This coincides with the periods of greatest recruitments of S. m. melanotheron in Lake Ahémé (Niyonkuru, 2007) and Lake Toho (Lederoun et al., 2015), and from Sarotherodon galilaeus galilaeus in Lake Doukon and Lake Togbadji (Lederoun et al., 2016). Our observations are consistent with those of Pauly (1980), who found that tropical fish species mostly exhibit double recruitment pulses. The two peaks of recruitment are observed during the rainy period and, particularly, during the period of flooding of the plains in August-September in southern Benin. This suggests that an abundance of food during the rainy seasons favors juveniles' recruitment into the population. The high proportions of individuals in the advanced gonad maturation stages (stages 3 and 4) were observed during these periods that Koné (2000) considers as the periods during which trophic conditions become favorable for young fish's growth.
The decrease in the size of a population can be attributed to two causes: natural death (M) (disease, predation, etc.) and fishing pressure (F). For this work, the mortality study was based on concepts defined by Beverton and Holt (1957) and Gulland (1963). Once the value of M was determined, the M/K ratio is often used to examine the accuracy of growth parameters (Chakraborty, 2001) and for a rapid diagnosis of fisheries (Pauly and Moreau, 1997). In the first case, it is supposed to be constant for a group of species or closely related families or taxa (Chakraborty, 2001). It should generally fall within the range of 1.12–2.5 (Beverton and Holt, 1959). In the second case, it is strictly greater than 2 for populations with a low longevity (Pauly and Moreau, 1997). The M/K ratio of 1.98 obtained in this study reflects the reliability of M and K, an excellent environmental state (Beverton and Holt, 1959), and a long-lived population (Pauly and Moreau, 1997). The M/K ratio of Lake Nokoué and the Porto-Novo Lagoon is similar to those obtained in Lake Doukon (2.06) and Lake Togbadji (1.99) for S. g. galilaeus (Lederoun et al., 2016) and in Lake Toho (2.36) for S. m. melanotheron (Lederoun et al., 2016).
In the studied species, the Z/K ratio was 2.86, expressing a predominance of mortality overgrowth, on the one hand, and a slight exploitation, on the other hand (Barry and Tegner, 1989). King and Etim (2004) have also reported that Z/K ratio of less than or approximately close to 2 indicates that the targeted species' population is lightly exploited for a mortality-dominated community. Besides, natural mortality was higher than fishing mortality. Therefore, demography is strongly influenced by natural mortality linked to predation, disease, or other causes not related to fishing. This higher natural mortality is associated with a higher growth coefficient (K). Chauvet (1988) then Pauly and Yañez-Arancibia (1994) indicated that lagoons' ecological conditions seem to have a positive influence on growth but not on the mortality coefficient.
The high values of M and K indicate that these fish have high biomass turnover rates. According to Sparre and Venema (1992), the high growth coefficients, the short asymptotic lengths, and the high natural mortality indicate that the fish matures early and has a short lifespan. According to these same authors, if the natural mortality rate M is high, the fish quickly reaches the age or the loss due to natural mortality exceeds the gain in biomass due to growth. Fishing mortality should be higher to catch fish before they die from natural causes. Fish populations are considered to be below optimal exploitation when fishing mortality is lower than natural mortality. High values of natural mortality (Z = 2.3 and M = 1.5) were observed by Niyonkuru et al. (2007) in Lake Nokoué and by Lederoun et al. (2015) in Lake Toho (Z = 1.95 and M = 1.37). Even if the use of landing nets was mentioned in Lake Toho, the high proportion of predators (See Vodougnon et al., 2018) in Lake Nokoué and the Porto-Novo Lagoon is mostly responsible for the high value of natural mortality recorded. The low fishing mortality observed is partly linked to the fact that the catch also involves other species enjoyed by the community and abundant in catches such as Coptodon guineensis, Chrysichthys nigrodigitatus, Ethmalosa fimbriata, Pellonula leonensis, Neochelon falcipinnis.
The size of the first catch (Lc) is, by definition, the size at which the catching gear selects 50% of the fish. Thus, it is estimated that fish within this size is in the unexploited phase, and, from this, they suddenly become exposed to full exploitation. In this study, the size at first capture (Lc = 9.2 cm) is greater than or equal to the size at first maturity (8.9 and 9.2 cm TL, respectively for males and females), supporting the earlier assertion that the stock of S. m. melanotheron in the Lake Nokoué and Porto-Novo Lagoon complex are not overexploited. In the studied site, about 79.7% of the caught fishes (Fig. 2) are >9.2 cm TL (L50 of females), which means that many individuals of S. m. melanotheron are captured after reaching the matured stage and adding to the biomass of the population.
Two assumptions and data are used to calculate the yield and the relative biomass per recruit: (1) the selection of type “knife-edge,” which excludes that fish of sizes lower than Lc are not caught by the gear and (2) the “ogive selection” which assumes that the probability of catching any fish depends on its length. This size is then obtained from the selectivity curve of the net (Sparre et al., 1989). From this size the fish become vulnerable to nets. In this study, we have used the second method because it seems logical that the probability of the fish being caught by a given gear depends on its size.
As far as the level of exploitation is concerned, the current exploitation rate estimated approximately at 0.31 was substantially equal to the optimal rate of 0.325 obtained with the relative yield curve per recruit Y ′/R. This result highlighted that the S. m. melanotheron population is currently at an optimum level of exploitation (Beverton and Holt, 1957; Lederoun et al., 2016). This situation is similar to the one mentioned on Lake Nokoué by Niyonkuru et al. (2007) but is contrary to what one observed in Lake Toho where species is strongly overexploited (Lederoun et al., 2015). The use of acadjas might explain the reasonable current exploitation rate recorded in this complex. But as reported by Lederoun et al. (2016) for Lake Doukon and Lake Togbadji, the rational exploitation of species could easily switch to overexploitation in the future if sensitization sessions advocating sustainable fishing methods are not organized. The government currently commissions a new study on reorganizing the acadjas in the complex and removing prohibited gear, which reassures the fishing pressure in the future. In southern Benin, fish is the primary protein source, and this complex is the leading supplier.
Growth parameters (L∞ = Asymptotic length, to = Theoretical age at length zero, tmax = Longevity, Φ ′ = Growth performance index, and K= Growth coefficient), mortality estimates (Z = Total mortality, M = Natural mortality, and F = Fishing mortality), and exploitation rates (E) from different fishing areas in West Africa, including the estimates of the present study.
5 Conclusion
The lengths of first capture and first sexual maturity observed during this study, the Z/K ratio, and the current exploitation rate are indicators of rational exploitation of S. m. melanotheron in the Lake Nokoué and Porto-Novo Lagoon complex. Compared to the known situation in most lakes in southern Benin in which the species are overexploited, it seems here that the number of acadjas and the area they occupy considerably limit the overfishing of the species. This could be explained since the exploitation generally takes place at least a year after the installation of the branches, allowing the fish to reach the size of maturity and contribute to the exploitable biomass. However, the reorganization of these sedentary fisheries is still essential to avoid conflicts between fishermen. We also recommend reducing the fishing effort from March to April and August, when juvenile fishes' recruitment is very intense.
Supplementary Material
Fig. S1. Length-converted catch curve for Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex. Solid dots are those used in calculating the parameters of the straight line, the slope of which (with sign changed) is an estimate of Z. Open dots represent fish not fully selected by the gear used in the fishery, and grey dots are those not used in mortality estimation. Access Supplementary Material
Acknowledgements
The authors sincerely thank the fishermen who work in the study area, especially Zinsou Mehinto (Lake Nokoué), Philippe Zannou, and Jacob Kounasso (Porto-Novo Lagoon), for their participation in data collection, Tobias Musschoot (FishBase, Royal Museum for Central Africa) for the help with Figures 2–8. We are grateful to Dr. Maarten Van Steenberge (MRAC; KU Leuven, Belgique) for insightful comments on an earlier version of the manuscript. We also thank the NWO-WOTRO Netherlands Organisation for Scientific Research Wotro Science for Global Development, which funded this project within the framework of Improving the Resilience of Continental Fishermen Communities and Aquatic Systems with regard to Overfishing and Water Quality Degradation in Benin.
References
- Abowei JFN, Davies OA, Eli AA. 2009. Study of the Lengthweight relationship and condition factor of five fish species from Nkoro River, Niger Delta, Nigeria. Res J Biol Sci 1: 94–98. [Google Scholar]
- Abowei JFN, Hart AI. 2007. Size, composition, age, growth, mortality and exploitation rate of Chysichthys nigrodigitatus from Nun River, Niger Delta, Nigeria. Afr J Appl Zool Envir Biol 9: 44–50. [Google Scholar]
- Abowei JFN, Hart AI. 2009. Some morphormetric parameters of ten finfish species from the Lower Nun River, Niger Delta, Nigeria. Res J Biol Sci 4: 282–288. [Google Scholar]
- Adjibade KN, Adite A, Arame H, Imorou RS, Sonon SP. 2019. Biodiversity and community structure of mormyridae (Pisces: Teleostei: Osteoglossiformes) from Niger River in Northern Benin: threats, conservation and valorization perspectives. Int J Sci 8: 106–116. [Google Scholar]
- Aglinglo C. Production de poissons dans les acadjas du lac Nokoué et de la lagune de Porto: Novo en République du Bénin, Mémoire de DES. Université de Liège, Belgique, 1998. [Google Scholar]
- Ahouansou Montcho S. Diversité et Exploitation des poissons de la rivière Pendjari (Bénin, Afrique de l'Ouest), Thèse de Doctorat. Université d'Abomey-Calavi, Bénin, 2011. [Google Scholar]
- Akintola SL, Anetekjai MA, Fakoya KA, Adewolu MA. 2010. Seasonal distribution and aspects of biology of Genus Macrobrachium from Badagry Creek, Nigeria. Nig J Fish 7: 16–24. [Google Scholar]
- Alhassan EH, Armah AK. 2011. Population dynamics of the African River Prawn, Macrobrachium vollenhovenii, in Dawhenya Impoundment. Turk J Fish Aquat Sci 11: 113–119. [CrossRef] [Google Scholar]
- Barry JP, Tegner MJ. 1989. Inferring demographic processes from size-frequency distributions: simple models indicate specific patterns of growth and mortality. Fish B-NOAA 88: 13–19. [Google Scholar]
- Bédia AT, N'doua Etilé R, Bi GG, Essetchi Kouamelan P, N'douba V. 2017. Paramètres de croissance et d'exploitation de Chrysichthys nigrodigitatus (Lecepede, 1803) (Siluriformes, Bagridae) dans une lagune tropicale: lagune Ebrié (Secteur I: lagune Potou, Côte d'Ivoire). Tropicultura 35: 253–261. [Google Scholar]
- Beverton RJH, Holt SJ. On the Dynamics of Exploited Fish Populations. Chapman and Hall, London, 1957, 538 p. [Google Scholar]
- Beverton RJH, Holt SJ. Manual of methods for fish stock assessment: part II. tables of yield function. FAO Fisheries Technical Paper No. 38. Food and Agriculture Organization, Rome, 1966, 67p. [Google Scholar]
- Branstetter S. 1987. Age and growth estimates for blacktip, Carcharhinus limbatus, and spinner Carcharhinus brevipinna, sharks from the northwestern Gulf of Mexico. Copeia 4: 964–974. [CrossRef] [Google Scholar]
- Brown-Peterson NJ, Wyanski DM, Saborido-Rey F, Macewicz BJ, Lowerre-Barbieri SK. 2011. A standardized terminology for describing reproductive development in fishes. Mar Coast Fish 3: 52–70. [Google Scholar]
- Chakraborty SK. 2001. Growth studies of sciaenids from Mumbai waters using the Bhattacharya Method. Naga, ICLARM Quart 24: 40–41. [Google Scholar]
- Chauvet C. Manuel sur l'amenagement des peches dans les lagunes cotieres : la bordigue mediterraneenne. FAO Fisheries Technical Paper, 1988, 290, 77p. [Google Scholar]
- Fagade SO. Age determination in Tilapia melanotheron (Rüppel) in the Lagos Lagoon, Lagos, Nigeria, with a discussion of the environmental and physiological basis of growth markings in the tropics, in Bagenal, TB (Ed.). The Ageing of Fish. Unwin Brothers Ltd, Old Woking, Surrey, England, 1974, pp. 71– 77. [Google Scholar]
- Fröese R. 2004. Keep it simple: three indicators to deal with overfishing. Fish Fish 5: 86–91. [CrossRef] [Google Scholar]
- Gaamour A. La Sardinelle ronde (Sardinella aurita Valenciennes, 1847) dans les eaux tunisiennes: reproduction, croissance et pêche dans la région du cap Bon, Thèse de Doctorat. Université de Bretagne occidentale, France, 1999. [Google Scholar]
- Gayanilo FC, Jr, Sparre P, Pauly D. FAO-ICLARM stock assessment tools (FiSAT). User's Guide. FAO Computerised Information Series (Fisheries) No. 8. Food and Agriculture Organization, Rome, 1996, 126p. [Google Scholar]
- Gayanilo FC, Sparre P, Pauly D. FAO-ICLARM population assessment tools II (FiSAT II). Revised. User's guide. Computerized Information Series (Fisheries). No. 8. Revised version. FAO, Rome, 2005, 168p. [Google Scholar]
- Gnohossou MP. La faune benthique d'une Lagune Ouest Africaine (le Lac Nokoué au Bénin), diversité, abondance, variations temporelles et spatiales, place dans la chaîne trophique, Thèse de doctorat. Ecole Nationale Superieure Agronomique de Toulouse, France, 2006. [Google Scholar]
- Gulland JA. 1963. The estimation of fishing mortality from tagging experiments. Int Comm Northwest Atl Fish Spec Publ 4: 218–227. [Google Scholar]
- Hilborn R, Walters CJ. Quantitative fisheries stock assessment-choice, dynamics and uncertainty. Chapman and Hall, New York, 1992, 570 p. [Google Scholar]
- Imorou RS, Adite A, Arame H, Chikou A, Adjibade NK, Sonon SP. 2019. Ichthyofauna of Okpara Stream, a Tributary of Oueme River, Benin, West-Africa. Int J Sci 8: 53–66. [Google Scholar]
- King M. 1995. Fisheries Biology: Assessment and Management. Fishing News Books, Oxford. [Google Scholar]
- King RP, Etim L. 2004. Reproduction, growth, mortality and yield of Tilapia mariae Boulenger 1899 (Cichlidae) in a Nigerian rainforest wetland stream. J Appl Ichthyol 20: 502–510. [CrossRef] [Google Scholar]
- Koné T. Régime alimentaire et reproduction d'un Tilapia lagunaire (Sarotherodon melanotheron melanotheron Rüppel, 1852) dans la rivière Bia et le lac de barrage d'Ayamé, Côte d'Ivoire), Thèse de Doctorat. Katholieke Universiteit, Leuven, 2000. [Google Scholar]
- Konan KF, Ouattara A, Ouattara M, Gourène G. 2007. Weight-length relationship of 57 fish species of the coastal rivers in South-Eastern of Ivory-Coast. Ribarstvo 65: 49–60. [Google Scholar]
- Lalèyè PA. Ecologie comparée de deux espèces de Chrysichthys, Poissons siluriformes (Claroteidae) du complexe lagunaire lac Nokouélagune de Porto-Novo au Bénin (Thèse de Doctorat), Université de Liège, Belgique, 1995. [Google Scholar]
- Lalèyè PA. Acadja fisheries enhancement systems in Benin: their productivity and environmental impacts, in Abban, EK, Casal, CMV, Falk, TM, Pullin, RSV (Eds.). Biodiversity and Sustainable Use of Fish in the Coastal Zone. The World Fish Center: Penang (Malaysia); ICLARM proceedings of an international workshop 63. 2000, pp. 51–52. [Google Scholar]
- Lalèyè PA, Niyonkuru C, Moreau J, Teugels GG. 2003. Spatial and seasonal distribution of the ichthyofauna of Lake Nokoué, Benin, West Africa. Afr J Aquat Sci 28: 151–161. [CrossRef] [Google Scholar]
- Lalèyè PA, Chikou A, Philippart J-C, Teuguels G, Vandewalle P. 2004. Étude de la diversité ichtyologique du bassin du fleuve Ouémé au Bénin (Afrique de l'Ouest). Cybium 28: 329–339. [Google Scholar]
- Le Cren ED. 1951. The length-weight relationship and seasonal cycle in gonadal weight and condition in the perch (Perca fluviatilis). J Anim Ecol 20: 201–219. [CrossRef] [Google Scholar]
- Lederoun D. Ichtyofaune du bassin du fleuve Mono (Bénin et Togo), diversité, écologie et impacts anthrophiques, Thèse de Doctorat. Université de Liège, Belgique, 2005. [Google Scholar]
- Lederoun D, Chikou A, Vreven E, Snoeks J, Moreau J, Vandewalle P, Lalèyè P. 2015. Population parameters and exploitation rate of Sarotherodon melanotheron melanotheron rüppell, 1852 (Cichlidae) in Lake Toho, Benin. JBES 6: 259–271. [Google Scholar]
- Lederoun D, Lalèyè KR, Boni AR, Amoussou G, Vodougnon H, Adjibogoun H, Lalèyè PA. 2018b. Length-weight and length-length relationships of some of the most abundant species in the fish catches of Lake Nokoué and Porto-Novo Lagoon (Benin, West Africa). Lake Reserv, 00: 1–7. [Google Scholar]
- Lederoun D, Snoeks J, Lalèyè P, Vandewalle P, Vreven E. 2018a. An updated checklist of the ichthyofauna of the Mono River basin (Benin and Togo: West Africa). Ichthyol Explor Fres 28: 135–156. [Google Scholar]
- Lederoun D, Vandewalle P, Brahim AA, Moreau J, Lalèyè PA. 2016. Population parameters and exploitation rate of Sarotherodon galilaeus galilaeus (Cichlidae) in Lakes Doukon and Togbadji, Benin. Afr J Aquat Sci 41: 151–160. [CrossRef] [Google Scholar]
- Leonardis I, Sinis A. 1998. Reproductive strategy of Aphanius fasciatus Nardo, 1827 (Pisces: Cyprinodontidae) in the Mesolongi and Etolikon lagoons (W. Greece). Fish Res 35: 171–181. [CrossRef] [Google Scholar]
- MacDonald PDM, Pitcher TJ. 1979. Age-groups from size-frequency data: a versatile and efficient method of analysing distribution mixtures. J Fish Res Board Can 36: 987–1001. [CrossRef] [Google Scholar]
- Montchowui E, Chikou A, Ovidio M, Lalèyè P, Poncin P. 2011. Population dynamics of Labeo senegalensis Valenciennes 1842 (Pisces: Cyprinidae) in the Oueme River, Benin. J Fish Int 6: 52–58. [Google Scholar]
- Montchowui E, Lalèyè P, Moreau J, Philippart J-C, Poncin P. 2009. Population parameters of African carp: Labeo parvus Boulenger, 1902 (Pisces: Cyprinidae) in the Ouémé River in Benin (West Africa). North-West J ZooL 5: 26–33. [Google Scholar]
- Montchowui E, Niyonkuru C, Ahouansou Montcho S, Chikou A, Lalèyè P. 2007. L'ichtyofaune de la rivière Hlan au Bénin (Afrique de l'Ouest). Cybium 31: 163–166. [Google Scholar]
- Moritz T. 2010. Fishes of Iguidi River: a small forest stream in South-East Benin. Ichthyol Explor Fres, 21: 9–26. [Google Scholar]
- Moritz T, Lalèyè PA. 2019. Fishes of the Pendjari National Park (Benin, West Africa). J Fish Biol 18: 1–57. [Google Scholar]
- Moritz T, Laleye PA, Koba G, Linsenmair KE. 2006. An annotated list of fish from the River Niger at Malanville, Benin, with notes on the local fisheries. VGI 5: 95–110. [Google Scholar]
- Niyonkuru C. Etude comparée de l'exploitation et de la démographie des poissons Cichlidés dans les lacs Nokoué et Ahémé au Bénin, Thèse de Doctorat. Université d'Abomey-Calavi, Bénin, 2007. [Google Scholar]
- Niyonkuru C, Lalèyè PA. 2010. Impact of acadja fisheries on fish assemblages in Lake Nokoué, Benin, West Africa. Knowl Manag Aquat Ec 399: 05. [CrossRef] [Google Scholar]
- Niyonkuru C, Lalèyè P, Villanueva MC, Moreau J. 2007. Population parameters of main fish species of Lake Nokoué (West Africa). J Afr Zool Special issue: 149–155. [Google Scholar]
- Panfili J, Mbow A, Durand JD, Diop K, Diouf K, Thior D, Ndiaye P, Laë R. 2004. Influence on the life-history traits of the West African black-chinned tilapia (Sarotherodon melanotheron): comparison between the Gambia and the Saloum estuaries. Aquat Living Resour. 17: 65–74. [CrossRef] [EDP Sciences] [Google Scholar]
- Pauly D. 1979. Theory and management of tropical multispecies stocks: a review with emphasis on the southeast Asian demersal fisheries. ICLARM Sud Rev 1: 35. [Google Scholar]
- Pauly D. 1980. On the interrelationships between natural mortality, growth parameters and mean environmental temperature in 175 fish stocks. J Cons Int Explor Mer 39: 175–192. [CrossRef] [Google Scholar]
- Pauly D. 1994. Quantitative analysis of published data on the growth, metabolism, food consumption, and related features of the red-bellied piranha, Serrasalmus nattereri (Characidae). Environ Biol Fish 41: 423–437. [CrossRef] [Google Scholar]
- Pauly D, Moreau J. Méthodes Pour L'évaluation des Ressources Halieutiques. Cépaduès Editions: Toulouse, France, 1997, 288p. [Google Scholar]
- Pauly D, Munro JL. 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte 2: 21–22. [Google Scholar]
- Pauly D, Yañez-Arancibia A. Fisheries in coastal lagoons, in Kjerfve, B (Ed.). Coastal Lagoon Processes. Elsevier Oceanography Series, 1994, 60. pp. 377–399. [CrossRef] [Google Scholar]
- Rjeibi O, Gaamour A, Missaoui H. 2011. Etude de la croissance de la langouste rouge, Palinurus elephas dans les eaux tunisiennes. Bull Inst Natn Scien Tech Mer de Salammbô 38: 41–54. [Google Scholar]
- Saila SB, Recksiek CW, Prager MH. 1988. Basic fishery science programs: a compendium of microcomputer programs and manual of operation. Fish Sci Aquacul 18: 1–230. [Google Scholar]
- Sarmento H, Isumbisho M, Stenuite S, Darchambeau F, Bruno L, Descy J-P. 2009. Phytoplankton ecology of Lake Kivu (eastern Africa): biomass, production and elemental ratios. Verh Internat Verein Limnol 30: 709–713. [Google Scholar]
- Sparre P, Ursin E, Venema S. Introduction to tropical fish stock species assessment. Part 1-Manual FAO. Fisheries technical Paper 306/1, Rome, 1989, 337p. [Google Scholar]
- Sparre P, Venema S.C. Introduction to tropical fish stock assessment. Part 1. Manual, FAO Fisheries technical Paper N° 306/1. FAO, Rome, 1992, 376p. [Google Scholar]
- Sokal R, Rohlf F. Introduction to Biostatistics (2nd edn). WH Freeman, New York, 1987, 363p. [Google Scholar]
- Tah L, Joanny T.G., N'Douba V.N., Kouassi J.N., Moreau J. 2010. Preliminary estimates of the population parameters of major fish species in Lake Ayamé I (Bia basin; Côte d'Ivoire). J Appl Ichthyol 26: 57–63. [CrossRef] [Google Scholar]
- Taylor CC. 1962. Growth equations with metabolic parameters. J Cons Int Explor Mer 27: 270–286. [CrossRef] [Google Scholar]
- Teugels GG, Thys van den Audenaerde DF, Cichlidae E, in Paugy D, Lévêque C, Teugels GG (Eds.). Faune des Poissons d'Eaux douces et saumâtres de l'Afrique de l'Ouest, Vol. 2. Collection Faune et Flore Tropicales n 40.Musée Royale de l'Afrique Centrale, Tervuren; Muséum National d'Histoire Naturelle, Paris;Institut de Recherche pour le Développement, Paris, 2003, pp. 520–600. [Google Scholar]
- Torrejon-Magallanes J. 2019. SizeMat: Estimate Size at Sexual Maturity. R package version 1.0. https://CRAN.R.project.org/package=sizeMat [Google Scholar]
- Tuck ID, Chapman CJ, Atkinson RJA. 1997. Population biology of the Norway lobster, Nephrops norvegicus (L.) in the Firth of Clyde, Scotland − I: growth and density. ICES J MAR SCI 54: 125–35. [CrossRef] [Google Scholar]
- Villanueva MCS. Biodiversite et relations trophiques dans quelques milieux estuariens et lagunaires de l'Afrique de l'Ouest: adaptations aux ressources environnementales, Thèse de Doctorat. Institut national Polytechnique de Toulouse, France, 2004. [Google Scholar]
- Vodougnon H, Lederoun D, Amoussou G, Adjibogoun D, Lalèyè P. 2018. Ecologic stress in fish population of Lake Nokoué and Porto-Novo Lagoon in Benin. IJFAS 6: 292–300. [Google Scholar]
- Wahle AR, Fogarty M. Growth and development: understanding and modelling variability in lobsters, in Phillips BF (Ed.). Lobsters: Biology, Management, Aquaculture and Fisheries. Blackwell, Oxford, 2006, pp. 1– 44. [Google Scholar]
- Welcomme RL. 1972. An evaluation of the acadja method of fishing as practised in the coastal lagoons of Dahomey (West Africa). J Fish Biol 4: 39–55. [CrossRef] [Google Scholar]
Cite this article as: Lederoun D, Amoussou G, Baglo IS, Adjibogoun H, Vodougnon H, Moreau J, Lalèyè PA. 2020. Growth, mortality and yield of Sarotherodon melanotheron melanotheron (Rüppell, 1852) in Lake Nokoué and the Porto-Novo Lagoon complex Benin, West Africa. Aquat. Living Resour. 33: 18
All Tables
Growth parameters (L∞ = Asymptotic length, to = Theoretical age at length zero, tmax = Longevity, Φ ′ = Growth performance index, and K= Growth coefficient), mortality estimates (Z = Total mortality, M = Natural mortality, and F = Fishing mortality), and exploitation rates (E) from different fishing areas in West Africa, including the estimates of the present study.
All Figures
|
Fig. 1 Study area, showing sampling sites (▲) and acadjas (■). |
| In the text | |
|
Fig. 2 Size frequencies for Sarotherodon melanotheron melanotheron (Cichlidae) in Lake Nokoué and Porto-Novo Lagoon complex (N = 1,745). |
| In the text | |
|
Fig. 3 Estimated size at first maturity for males (a) and females (b) of Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex. |
| In the text | |
|
Fig. 4 Length frequency data superimposed on the growth curve (Continuous line) of Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex (L∞ = 24.68 cm total length, K = 0.86 y−1). The bars represent the restructured length frequency data, where black bars indicate positive peaks and white bars represent negative peaks. |
| In the text | |
|
Fig. 5 Probability of capture of Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex estimated from the ascending axis of the catch curve. |
| In the text | |
|
Fig. 6 Relative yield-per-recruit and biomass-per-recruit curves for Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex using the selection ogive option. The three dashed right-angled lines correspond to E0.5, E0.1 and Emax, respectively. |
| In the text | |
|
Fig. 7 Recruitment pattern of Sarotherodon melanotheron melanotheron from Lake Nokoué and Porto-Novo Lagoon complex showing two recruitment peaks per year. |
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.